REPOSITÓRIO CIENTÍFICO
Dinos 101
132 espécies curadas a partir da literatura paleontológica. Onde viveram, quais ossos foram encontrados, e como nossa compreensão evoluiu.

EXPLORE POR PERÍODO



























































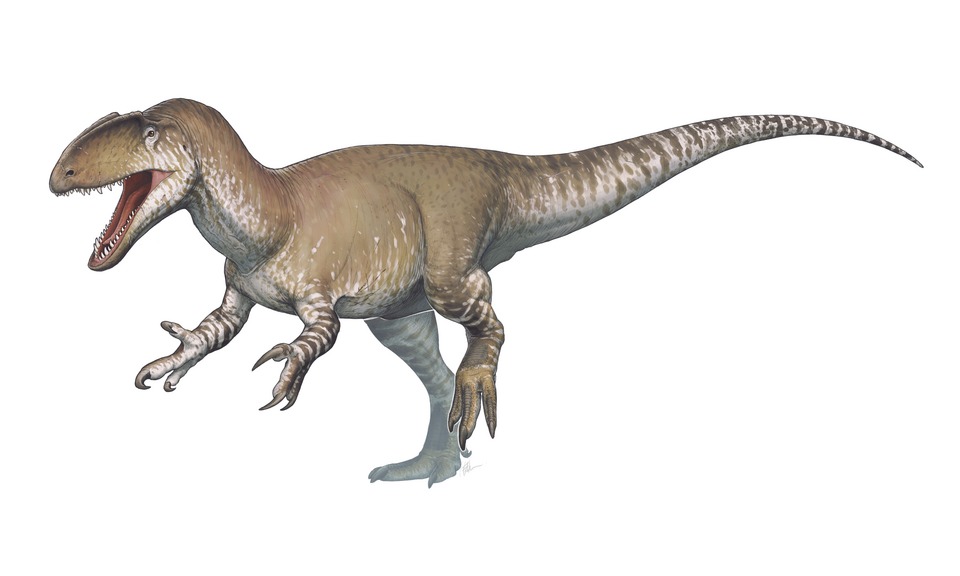
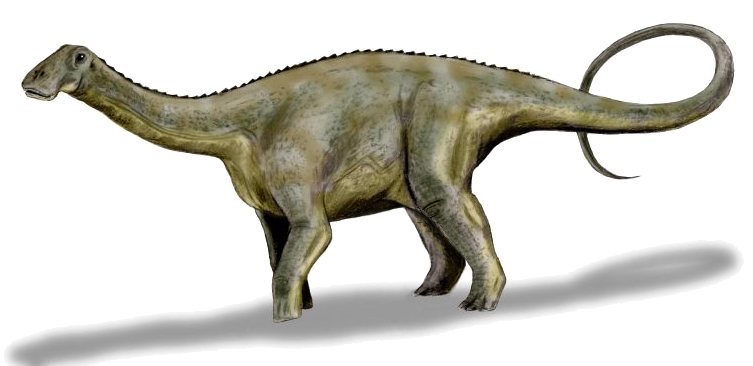

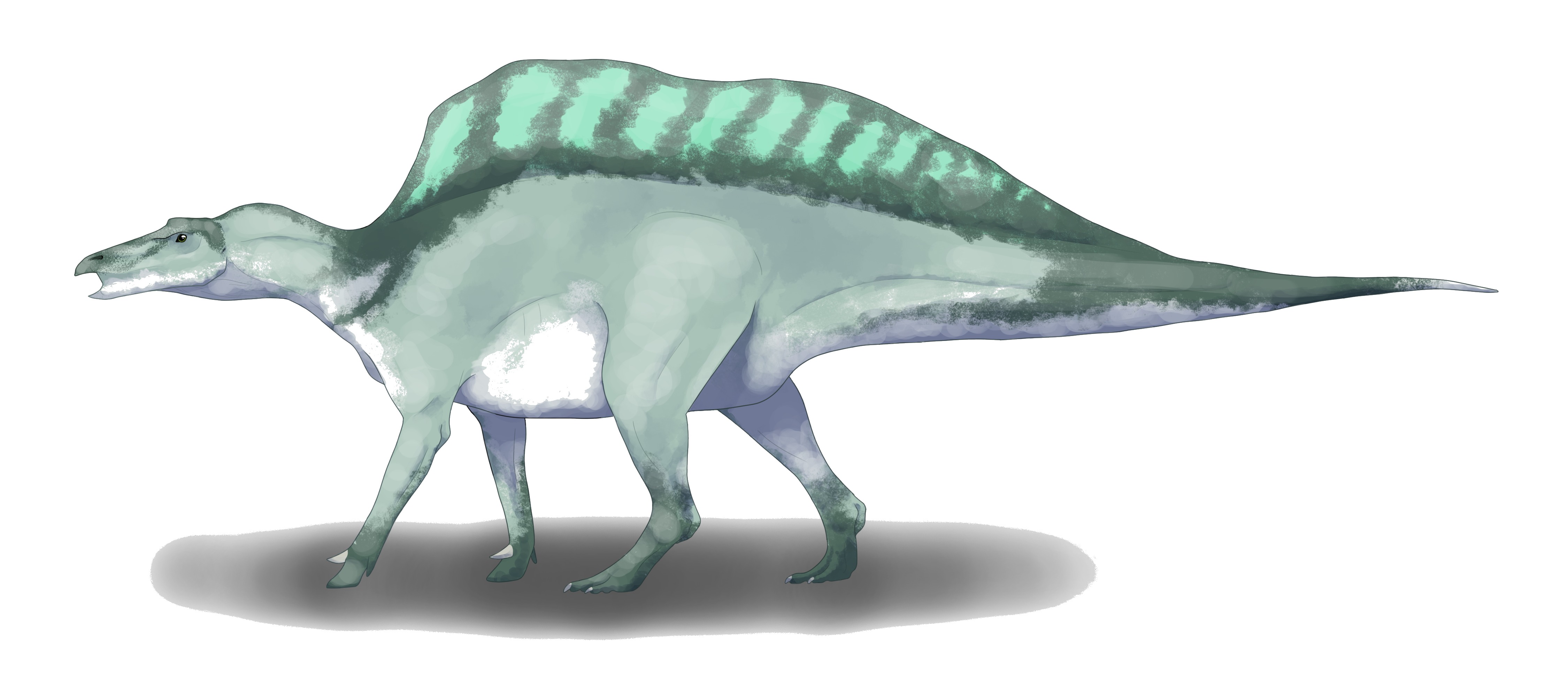







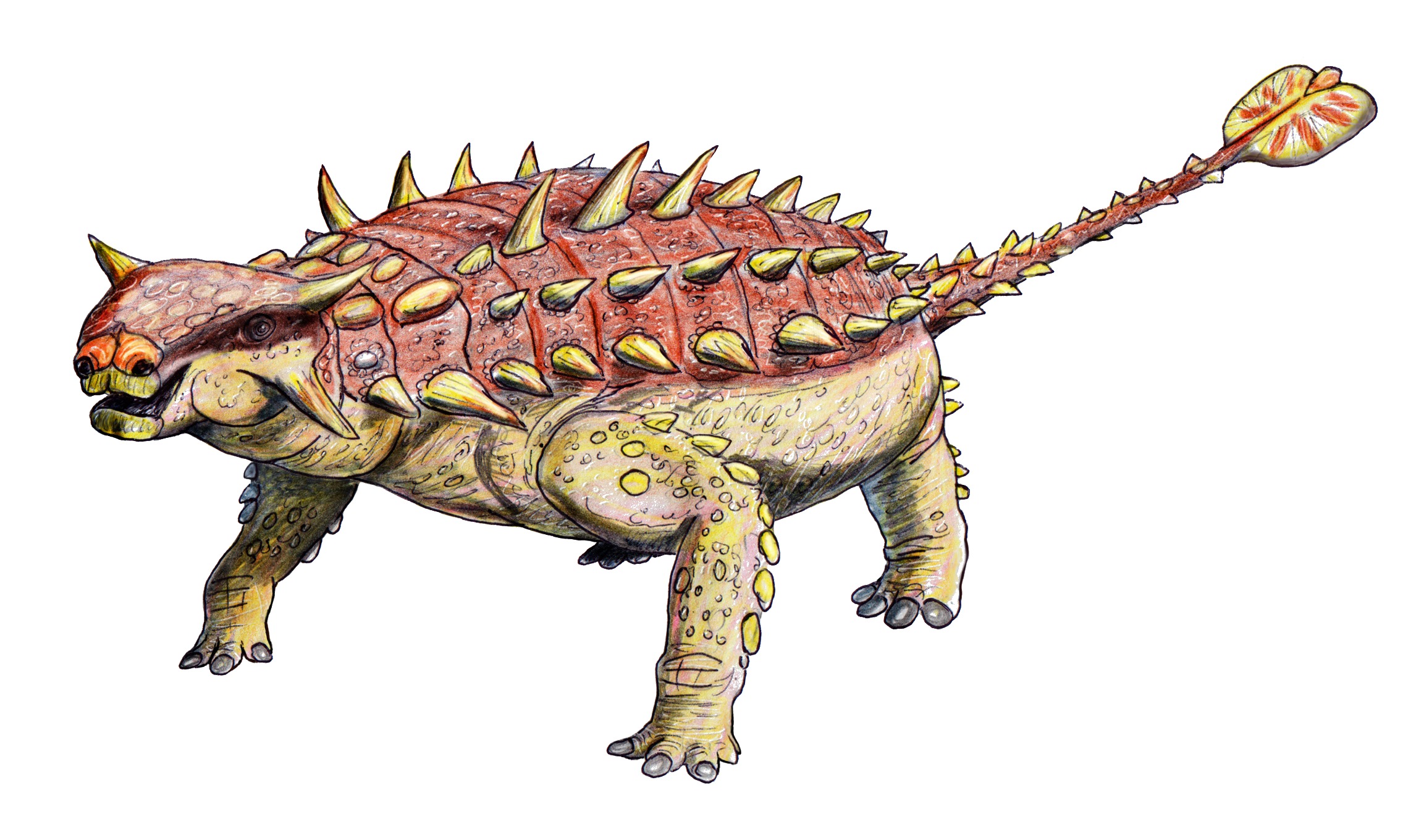

































Nenhuma espécie encontrada com esses filtros.
Mesozoico · Não-dinossauros
Outros Animais do Mesozoico
Não são dinossauros, mas dominaram os oceanos, o ar e parte dos continentes no mesmo período.
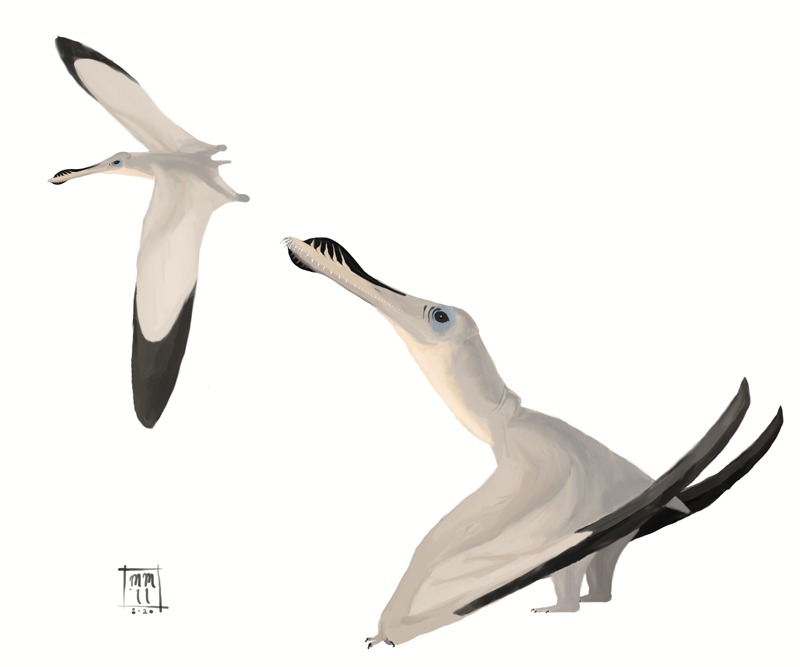
BR · 112–108 Ma
Anhanguera blittersdorffi
Anhanguera blittersdorffi
"Espírito maligno de Blittersdorff"
Anhanguera blittersdorffi é um pterossauro anhanguerídeo do Cretáceo Inferior do Brasil, descrito por Campos e Kellner em 1985 a partir de um crânio tridimensional extraordinariamente preservado. Com envergadura estimada entre 4 e 4,5 metros, era um predador aéreo especializado na captura de peixes, dotado de cristas premaxilares características, dentes coniformes curvos e mandíbula com forma de roseta. Proveniente dos nódulos calcários da Formação Romualdo, Bacia do Araripe, no Ceará, é um dos pterossauros brasileiros mais estudados e emblema da fauna cretácea do nordeste brasileiro.

US · 100–72 Ma
Tubarao-Ginsu
Cretoxyrhina mantelli
"Dente afiado do Cretáceo de Mantell"
Cretoxyrhina mantelli, popularmente conhecido como tubarão-Ginsu, foi um dos maiores e mais temidos predadores marinhos do Cretáceo tardio. Viveu há cerca de 100 a 72 milhões de anos no Mar Interior Ocidental, uma vasta via maritima que dividia a América do Norte. Com até 6,5 metros de comprimento e anatomia similar a de tubarões-mako modernos, predava mosassauros, plesiossauros, tartarugas marinhas e pterossauros. Vários esqueletos quase completos foram encontrados no Cretáceo do Kansas, tornando-o um dos tubarões extintos mais bem documentados da ciência.

DE · 157–137 Ma
Dakossauro
Dakosaurus maximus
"Lagarto-mordedor máximo"
Dakosaurus maximus foi o crocodiliformes marinho mais temível do Jurássico tardio. Pertencente à família Metriorhynchidae, distinguia-se de todos os outros membros do grupo por possuir dentes comprimidos lateralmente e serrilhados, convergentes com os de terópodes terrestres, indicando especialização em presas grandes. Sem armadura osteodermal e com membros transformados em nadadeiras e cauda bifurcada em forma de meia-lua, era totalmente adaptado à vida pelágica. Seus fósseis foram encontrados na Europa (França, Alemanha, Suíça, Polônia, Inglaterra) e na Argentina, onde o espécime D. andiniensis recebeu o apelido de 'Godzilla' pelos paleontólogos argentinos. Era o predador de topo dos mares jurássicos europeus.

US · 80.5–77 Ma
Elassossauro
Elasmosaurus platyurus
"Réptil de placa fina de cauda achatada"
Elasmosaurus platyurus foi um dos maiores e mais característicos plesiosaurídeos do Cretáceo Superior, famoso pelo pescoço extraordinariamente longo que representava mais da metade de seu comprimento total de cerca de 14 metros. Com 71 vértebras cervicais, ostenta o maior número de vértebras do pescoço de qualquer vertebrado conhecido. Não era um dinossauro, mas um réptil marinho do grupo Sauropterygia. Vivia no Mar Interior Ocidental que cobria o centro da América do Norte, alimentando-se de peixes e cefalópodes com seus dentes afiados. Tornou-se famoso por um erro histórico: quando Edward Drinker Cope o descreveu em 1868, montou o esqueleto com o crânio na ponta errada, posicionando-o na extremidade da cauda. O erro só foi corrigido em 1870, após a intervenção de Joseph Leidy.

DE · 183–174 Ma
Tubarao Hybodus
Hybodus hauffianus
"Tubarao giboso de Hauff"
Hybodus hauffianus é um dos tubarões extintos mais bem documentados do Mesozoico. Viveu no inicio do Jurássico, há cerca de 183 a 174 milhões de anos, nos mares rasos e quentes que cobriam a Europa central. Com até 2 metros de comprimento, possuía dois tipos de dentes: pontudos na frente para capturar presas escorregadias e achatados nas regiões posteriores para esmagar presas de concha dura. Especimes excepcionais preservados no Folhelho Posidonia da Alemanha revelam detalhes anatomicos raros, incluindo conteudo estomacal com rostra de belemniticos, confirmando sua dieta de cefalopodes.

AU · 115–100 Ma
Cronossauro
Kronosaurus queenslandicus
"Réptil de Kronos de Queensland"
Kronosaurus queenslandicus foi um dos maiores pliosaurídeos do Cretáceo Inferior, com comprimento estimado em 9 a 11 metros e crânio de 2,2 metros de comprimento, um dos maiores de qualquer réptil pré-histórico. Não era um dinossauro, mas um réptil marinho do clado Pliosauridae. Viveu no Mar de Eromanga que cobria o interior da Austrália durante o Aptiano-Albiano (~115-100 Ma). O apelido 'Plasterosaurus' reflete a controversa reconstrução de Harvard (MCZ 1285), onde oito vértebras extras de gesso foram adicionadas ao espécime, inflando o comprimento de 10,5 para 12,8 metros. Com dentes cônicos de até 7 centímetros, era o superpredador dominante de seu ambiente, capaz de atacar plesiosauros, tartarugas marinhas e grandes peixes.
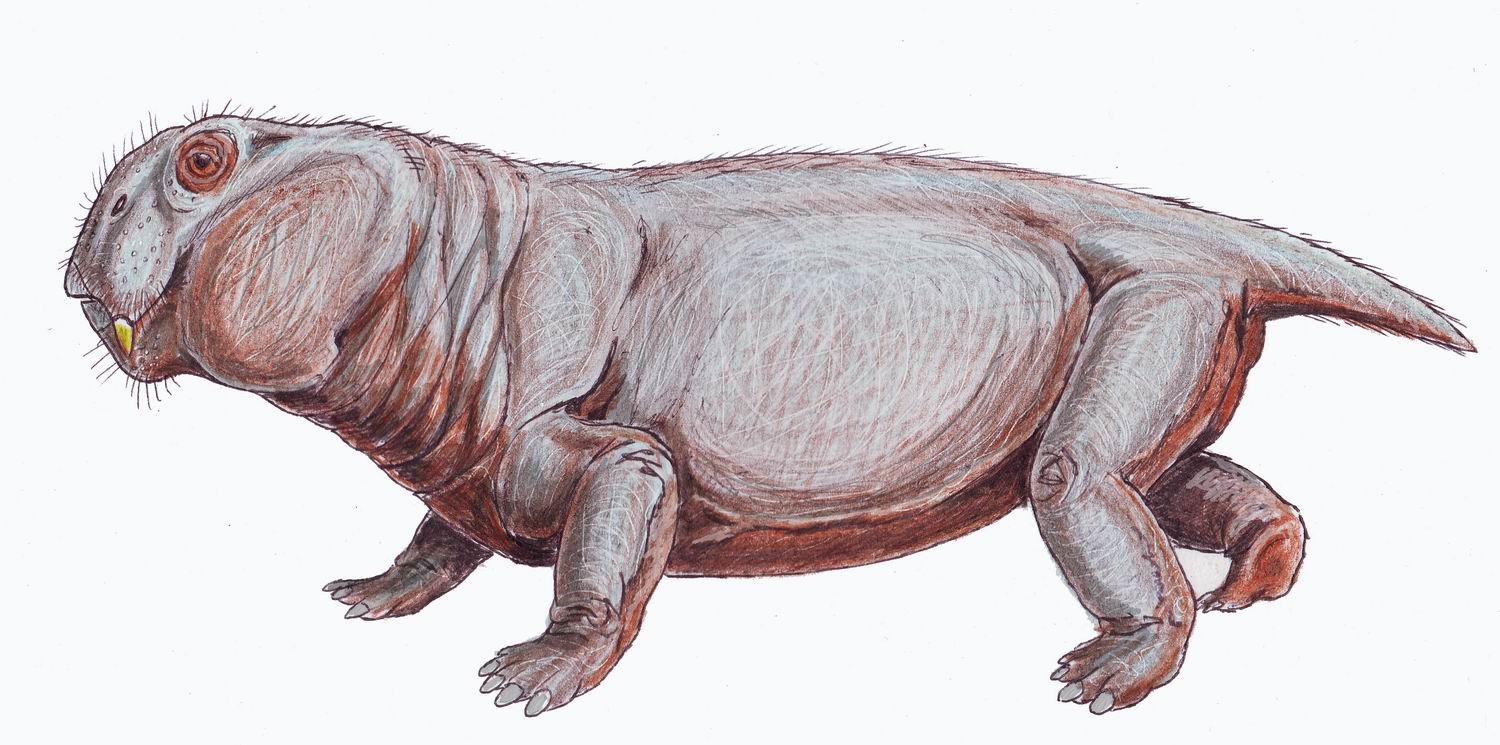
ZA · 252–249 Ma
Lystrossauro
Lystrosaurus murrayi
"Lagarto pá de Murray"
Lystrosaurus murrayi é um dicinodon (sinapsídeo não-mamífero) que sobreviveu à maior extinção em massa da história da vida na Terra, a catástrofe do final do Permiano, há cerca de 252 milhões de anos. No início do Triássico, chegou a constituir mais de 90% dos vertebrados terrestres, uma dominância sem paralelo na história dos tetrápodes. Seu crânio maciço portava um bico córneo para cortar vegetação e dois pequenos presas superiores. Com porte semelhante ao de um porco médio, Lystrosaurus murrayi tornou-se símbolo tanto da resiliência pós-extinção quanto da deriva continental, sendo encontrado na África do Sul, Antártica, Índia, China e Rússia.

NL · 82–66 Ma
Mosassauro
Mosasaurus hoffmannii
"Lagarto do Rio Mosa de Hoffmann"
O Mosasaurus hoffmannii foi o maior mosassauro conhecido e um dos maiores predadores marinhos de todos os tempos. Com até 13 metros de comprimento (estimativas recentes revisadas de valores anteriores de 17 metros) e peso estimado em 10 toneladas, dominava os oceanos do Cretáceo Superior. Não era um dinossauro, mas um réptil escamado (Squamata), parente próximo de monitores e cobras. Possuía mandíbula dupla articulada similar a das cobras, permitindo engolir presas grandes. Seus dentes robustos e cônicos eram adaptados para uma dieta generalista: peixes, tubarões, cefalópodes, tartarugas marinhas, aves e outros mosassauros. O primeiro fóssil de Mosasaurus, encontrado em Maastricht (Países Baixos) em 1764, foi um dos primeiros répteis marinhos gigantes descritos pela ciência, antes mesmo de Darwin. O crânio holótipo foi confiscado por soldados franceses durante o Cerco de Maastricht em 1794 e levado a Paris, onde Georges Cuvier o usou como prova de que espécies podiam se extinguir, conceito revolucionário na época.

GB · 165–150 Ma
Ictiossauro de Olhos Gigantes
Ophthalmosaurus icenicus
"Lagarto dos olhos (em grego: ophthalmos = olho, sauros = lagarto)"
Ophthalmosaurus icenicus foi um ictiossauro do Jurássico Médio-Superior, famoso pelos maiores olhos em proporção corporal de qualquer vertebrado conhecido: o anel escleral media até 23 centímetros de diâmetro externo. Encontrado principalmente na Formação Oxford Clay de Peterborough, Inglaterra, era um nadador oceânico ágil, com corpo hidrodinâmico de cerca de 6 metros e nadadeiras em forma de remo. Apesar de não ser um dinossauro, era estritamente contemporâneo a muitos deles. Seus olhos descomunais eram adaptados para mergulhos profundos em zonas mesopelágicas escuras, provavelmente em busca de lulas e cefalópodes. Harry Govier Seeley descreveu a espécie em 1874, e desde então tornou-se um dos ictiossauros mais bem documentados do Jurássico, com dezenas de espécimes excelentes preservados.

US · 228–201 Ma
Postosuchus
Postosuchus kirkpatricki
"Crocodilo de Post (cidade de Post, Texas)"
Postosuchus kirkpatricki foi um dos maiores predadores terrestres do Triássico Superior norte-americano. Arcossauro crurotarsal pertencente à família Rauisuchidae, ele não era um dinossauro, mas sim um parente distante dos crocodilos modernos. Com comprimento estimado entre 4 e 6 metros, possuía postura ereta, dentes serrilhados comprimidos lateralmente e crânio robusto. A locomoção era predominantemente bípede em adultos, com os membros anteriores progressivamente reduzidos com o crescimento. Viveu nos ambientes tropicais úmidos e semiáridos do Triássico tardio, convivendo com os primeiros dinossauros e os rincossauros que eventualmente substituiria.

US · 88–80 Ma
Pteranodonte
Pteranodon longiceps
"Asa desdentada de cabeça longa"
Pteranodon longiceps é o pterossauro mais estudado da história da paleontologia, com mais de 1.200 espécimes conhecidos. Voava sobre o Mar Interior Ocidental, uma via aquática rasa que cobria o centro da América do Norte no Cretáceo. Machos adultos atingiam envergadura de 5,6 a 7,6 metros, enquanto fêmeas eram menores, com aproximadamente 3,8 metros. A crista cefálica longa e voltada para trás era mais proeminente em machos. Apesar da aparência imponente, era um piscívoro especializado, capturando peixes na superfície do mar com o bico longo e sem dentes.

US · 68–66 Ma
Quetzalcoatlus northropi
Quetzalcoatlus northropi
"Serpente emplumada de Northrop"
Quetzalcoatlus northropi é o maior pterossauro conhecido e um dos maiores animais voadores da história da Terra. Com envergadura estimada em 10 a 11 metros, era tão largo quanto um avião monomotor. Vivia no Maastrichtiano tardio, há 68 a 66 milhões de anos, na Formação Javelina do Texas, na bacia do atual Parque Nacional Big Bend. Apesar do porte colossal, pesava apenas 150 a 250 kg graças aos ossos ocos com traves internas. Estudos biomecânicos demonstram que era capaz de voo ativo, decolando em salto quadrúpede com os membros anteriores. Em terra, caminhava em postura quadrúpede e caçava vertebrados pequenos à maneira de uma cegonha gigante.

CN · 125–123 Ma
Repenomamus
Repenomamus robustus
"Mamífero réptil robusto"
Repenomamus robustus foi um dos maiores mamíferos do Mesozoico e prova viva de que os mamíferos da Era dos Dinossauros não eram todos pequenos e inofensivos. Pertencente à ordem Eutriconodonta e à família Gobiconodontidae, viveu no Aptiano do Cretáceo Inferior em Liaoning, China, na mesma fauna do Microraptor gui. Com cerca de 50 cm de comprimento corporal e peso estimado em 4,5 kg, era tão grande quanto um gambá-da-Virgínia moderno. O achado mais notável é um espécime com ossos de um filhote juvenil de Psittacosaurus fossilizados no estômago, a única evidência direta de mamífero mesozoico predando dinossauros. A mandíbula robusta e os dentes heterodontes confirmam hábitos francamente carnívoros.

US · 221–205 Ma
Fitossauro-facão
Smilosuchus gregorii
"Crocodilo-faca de Gregory (homenagem ao geólogo Herbert E. Gregory)"
Smilosuchus gregorii foi um dos maiores predadores semiaquáticos do Triássico tardio da América do Norte. Com até 6 metros de comprimento, ocupava ecologicamente o mesmo nicho dos crocodilos modernos: emboscadas na margem de rios e lagos da Formação Chinle, no atual Arizona. Sua principal distinção em relação aos crocodilos verdadeiros está na posição das narinas: em vez de ficarem na ponta do focinho, as narinas dos fitossauros se abrem em uma protuberância entre os olhos. O crânio de S. gregorii pode ultrapassar 1,5 metro de comprimento, com dentes heterodônticos: grandes presas anteriores para impalar presas e dentes mais cortantes na região posterior para fatiar carne. A espécie foi descrita originalmente por Camp (1930) e transferida para o gênero Smilosuchus por Long e Murry (1995).

US · 100–72 Ma
Tubarão Corvo
Squalicorax falcatus
"Tubarão corvo falcado (pela curvatura dos dentes)"
Squalicorax falcatus foi um tubarão lamniforme de médio porte que habitou os mares rasos do Cretáceo Superior, incluindo o extenso Mar Interior Ocidental da América do Norte. Com cerca de 2,5 metros de comprimento, tinha o corpo fusiforme similar ao do tubarão-recife moderno, mas os dentes fortemente serrilhados lembravam os do tubarão-tigre atual. O gênero Squalicorax, popularmente chamado de tubarão-corvo, era um predador generalista e carniceiro oportunista. Evidências fósseis incluem dentes embutidos em ossos de hadrossaurídeos terrestres, mosassauros e tartarugas marinhas, revelando que se alimentava de carcaças arrastadas ao mar. A espécie S. falcatus é conhecida por esqueletos quase completos encontrados no Kansas, tornando-a uma das mais bem documentadas entre os tubarões mesozoicos.