Paleobiologia
Biologia dos Dinossauros
Como funcionavam os maiores animais terrestres da história? Do metabolismo ao comportamento social, das penas a inteligência: o que as evidências fósseis revelam sobre a biologia dos dinossauros.
Por que ficaram tão grandes?
O Brachiosaurus pesava 50 toneladas. O Patagotitan, descoberto em 2017, pode ter chegado a 70. Para entender como animais terrestres chegaram a esse tamanho, e preciso olhar para a anatomia.
🫁
Sistema respiratório com sacos aéreos
Passaros respiram de forma unidirecional: o ar passa pelos pulmões em uma só direção, sem nunca "voltar". Os dinossauros faziam o mesmo, usando sacos aéreos que funcionavam como foles. Isso extrai muito mais oxigênio por respiração do que o sistema bidirecional dos mamíferos, tornando corpos grandes energeticamente viáveis. Um corpo de 50 toneladas com pulmões de mamífero séria inviável: não conseguiria oxigenar os tecidos com eficiência suficiente.
🦴
Ossos ocos por dentro
As vértebras de Brachiosaurus eram até 60% ar. Os ossos dos saurópodes tinham câmaras internas conectadas aos sacos aéreos, mantendo estrutura rígida sem o peso correspondente. Um elefante africano de 6 toneladas tem ossos proporcionalmente mais densos do que um saurópode de 50 toneladas. Essa pneumatização óssea e uma das razões pelas quais dinossauros e pássaros compartilham a capacidade de crescer além dos limites dos mamíferos.
🌿
Engolir sem mastigar
Sauropodes não mastigavam. Arrancavam plantas e engoliam inteiro, deixando a fermentação no intestino fazer o trabalho. Uma cabeça pequena e leve no topo de um pescoço enorme consumia vegetação em grande volume sem desperdiçar energia mastigando. Quanto menor a cabeça, mais fácil sustentar o pescoço longo. E o oposto de um elefante, que tem cabeça enorme cheia de músculos mastigatórios e molares de alta coroa.
📈
Crescimento rápido como pássaros
Répteis crescem lentamente a vida toda. Passaros crescem rápido e param. Dinossauros cresciam como pássaros. Um saurópode jovem ganhava mais de 2 toneladas por ano no pico de crescimento e chegava ao tamanho adulto em 20 a 30 anos. Isso reduzia o tempo que o animal ficava vulnerável a predadores. Uma elefante demora 20 anos para atingir 5 toneladas. Dinossauros atingiam 10 vezes isso em tempo similar.
🌳
Abundancia de plantas no mundo quente
O CO₂ elevado do Mesozoico era combustível para as plantas. Florestas densas cobriam latitudes que hoje são desertos ou tundra. Mais biomassa vegetal significa que herbívoros gigantes conseguiam se alimentar sem esgotar o ambiente. Um saurópode precisava comer centenas de quilos de vegetação por dia. Só era possível porque havia comida em toda parte.
💨
O papel do oxigênio
O O₂ atmosférico subiu para cerca de 30% no Cretáceo tardio, contra 21% hoje. Isso ajudou, mas tem um contexto importante: os insetos gigantes do Carbonífero, 300 milhões de anos antes dos dinossauros, cresceram com O₂ em 35%. Eles eram grandes por razões completamente diferentes. Para os dinossauros, o sistema de sacos aéreos e o fator respiratório central. O nível de oxigênio foi um facilitador, não a causa.
Clima frio fez os dinossauros crescerem?
É comum ver a ideia de que resfriamentos climáticos no fim do Cretáceo empurraram os dinossauros para tamanhos gigantes, aplicando a regra de Bergmann (animais de sangue quente ficam maiores em climas frios para conservar calor). Um estudo de 2025 testou essa hipótese para tiranossauros com estatística rigorosa e chegou a uma resposta mais sutil.
O estudo mapeou três variáveis distintas sobre a mesma árvore de parentesco dos tiranossauroides: massa corporal, comprimento e temperatura média anual do local em que cada espécie viveu. O tempo geológico corre da esquerda (Jurássico, 200 milhões de anos) para a direita (fim do Cretáceo, 66 milhões de anos). As cores dos ramos indicam o valor estimado da variável naquele momento evolutivo: tons frios (roxo, azul) são valores baixos, tons quentes (amarelo, laranja, vermelho) são valores altos. Compare os três painéis abaixo.
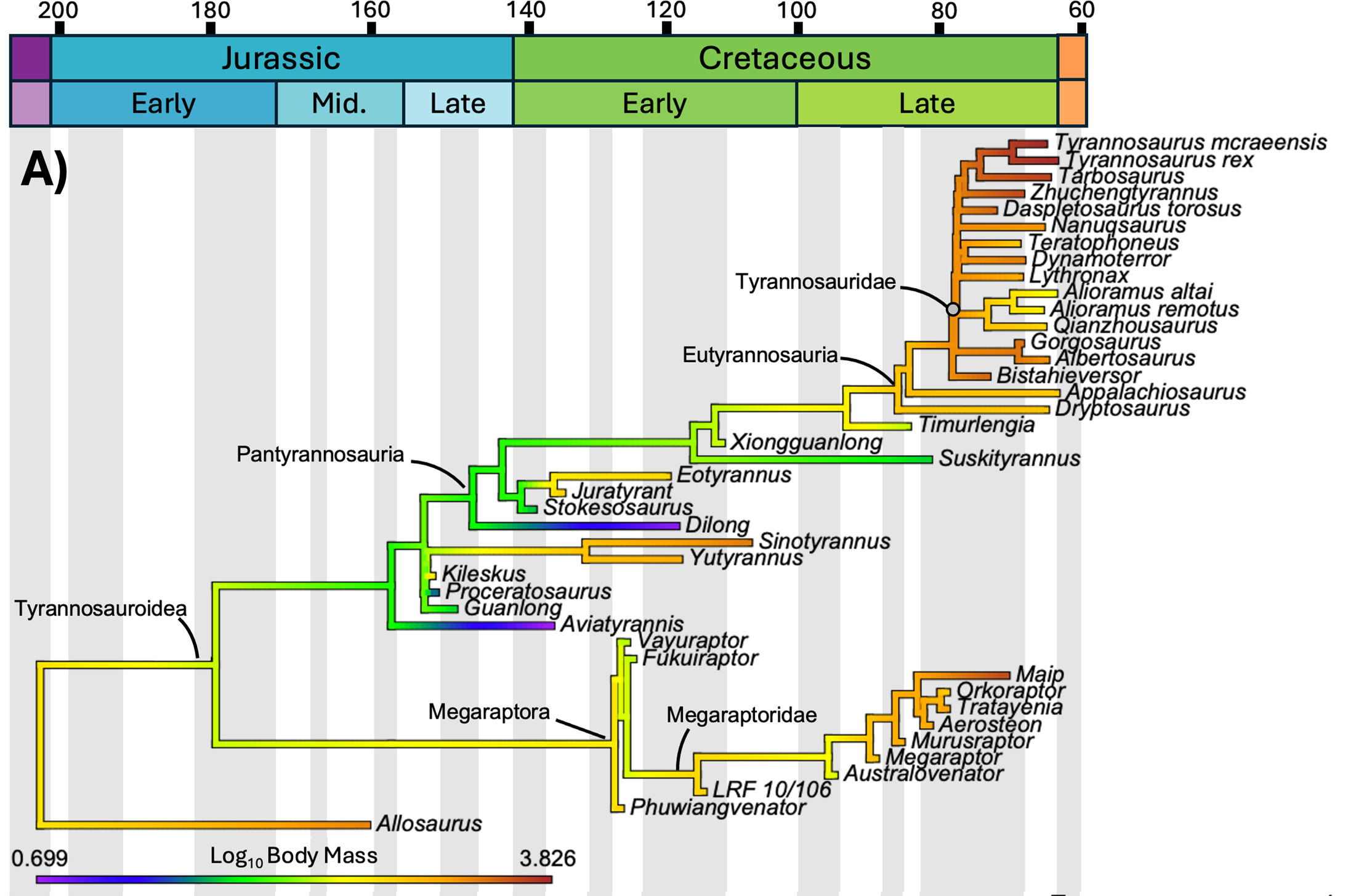
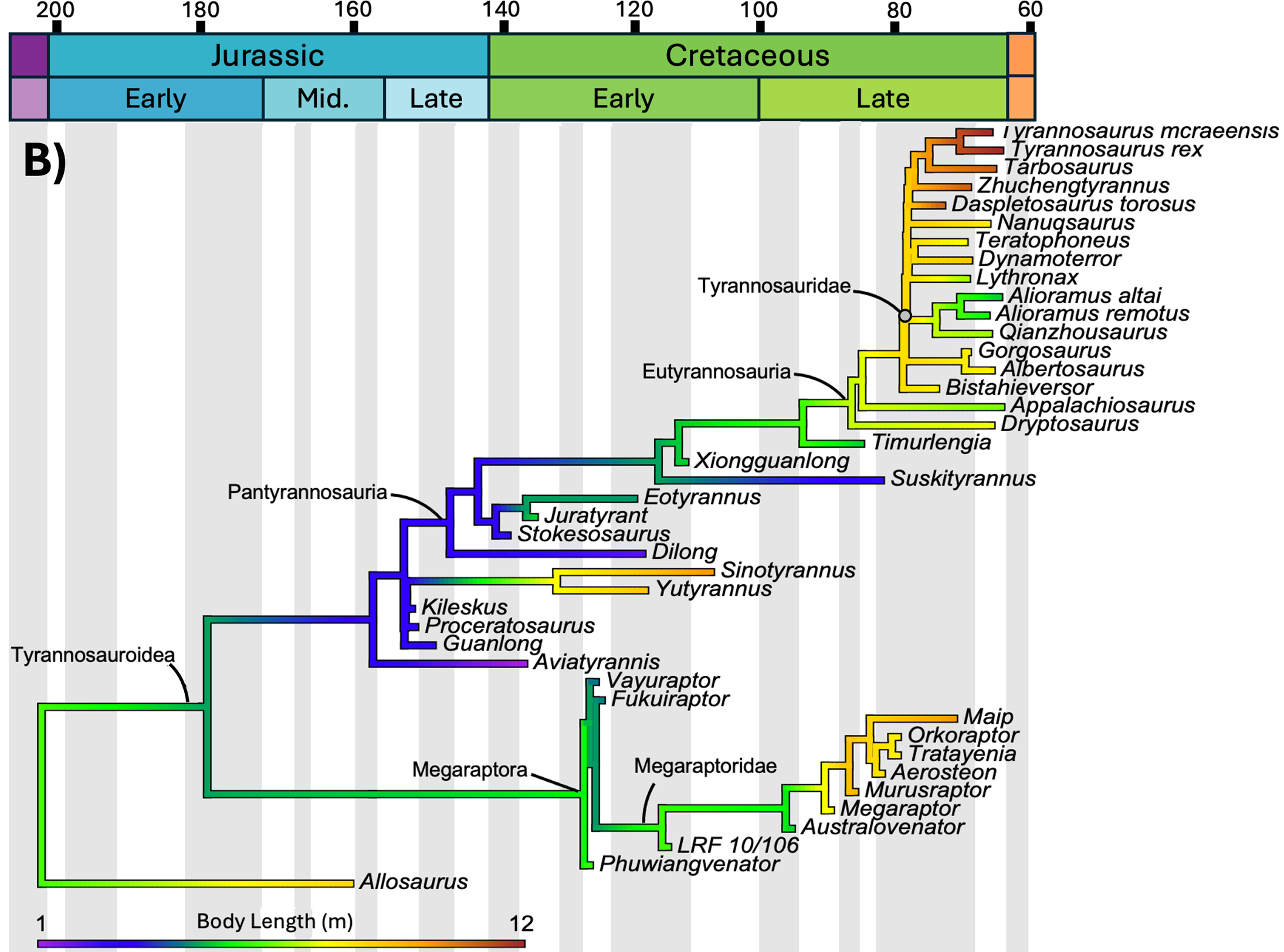
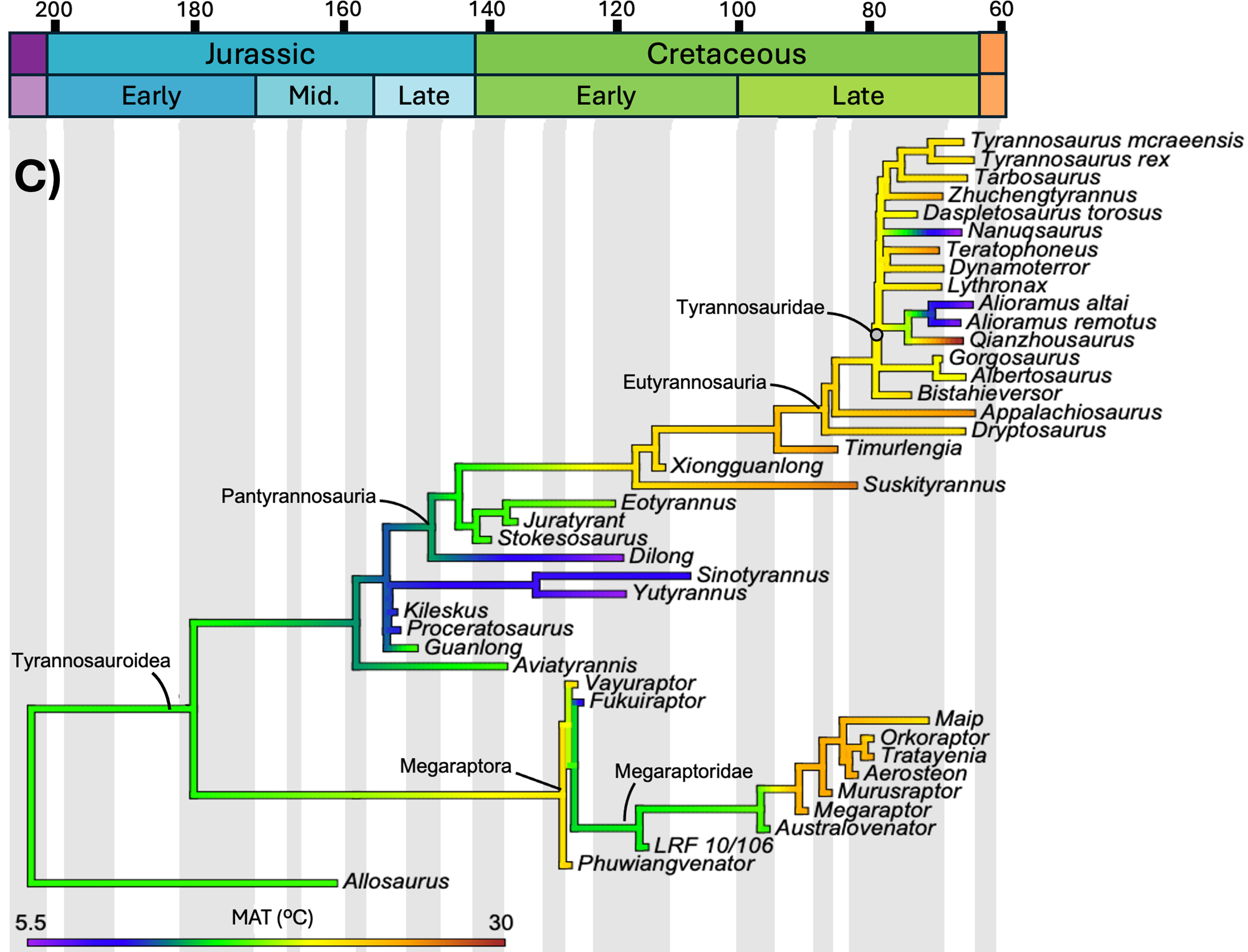
O teste estatístico
Morrison e colaboradores mapearam a evolução da massa corporal e do comprimento de cada tiranossauro conhecido contra a temperatura média anual (MAT) da época e da região em que viveu. A correlação entre temperatura e massa corporal foi estatisticamente não significativa: R² igual a 0,303 e p igual a 0,051, à beira do limiar de significância mas sem passar dele. Mesmo depois de aplicar logaritmo aos dados, o resultado não melhorou (R² igual a 0,293, p igual a 0,060).
Em português claro: a temperatura não prevê o tamanho corporal de um tiranossauro. Não existe relação direta do tipo "ficou mais frio, ficou maior".
Mas o estudo também mostra que os tiranossauros realmente ficaram gigantes em um momento específico: logo depois do chamado Máximo Térmico do Cretáceo, entre 90 e 95 milhões de anos atrás, quando as temperaturas globais começaram a cair em direção ao Campaniano e Maastrichtiano. A resposta dos autores para essa aparente contradição é que o clima atuou de forma indireta.
O resfriamento pós-Máximo Térmico contribuiu para a extinção dos carcarodontossaurídeos, o grupo de predadores apex que incluía o Giganotossauro, o Carcharodontosaurus e o Mapusaurus. Esses animais tinham 12 a 13 metros e dominavam o topo da cadeia alimentar na maior parte do mundo até o fim do Cenomaniano. Quando desapareceram, a vaga ecológica de "predador colossal de presas muito grandes" ficou aberta. Foi aí que os tiranossaurinos, até então carnívoros medianos de 5 a 7 metros, cresceram em poucos milhões de anos para os 12 metros do T. rex. Ou seja, o clima não fez o tiranossauro ficar maior diretamente; matou o concorrente, liberou o nicho.
Um detalhe curioso reforça o raciocínio: o Yutyrannus huali, um tiranossauro basal de 9 metros que viveu na China há 125 milhões de anos, em ambiente frio e de altitude elevada, já era gigante para a sua época. Os autores sugerem que a capacidade de alcançar grande porte em climas mais frios pode ser uma característica ancestral da linhagem, apenas realizada em escala quando o nicho permitiu. Mas ressalvam que o padrão não é uniforme no registro fóssil de tiranossauros e precisa de mais dados para ser confirmado.
Fonte: Morrison, C., Pittman, M., Nurgaliyev, B., & Salguero-Gómez, R. (2025). Rise of the king: Gondwanan origins and evolution of megaraptoran dinosaurs. Royal Society Open Science, 12(5), 242238. doi.org/10.1098/rsos.242238
Clima quente fez os dinossauros crescerem?
A pergunta simétrica também precisa ser feita. Se o frio não faz o dinossauro crescer, o calor faz? O Cretáceo médio foi o período mais quente de toda a Era Mesozoica e também abrigou os maiores animais terrestres que já pisaram no planeta. Há conexão de causa e efeito, ou coincidência?
O mundo em chamas (literalmente)
O Máximo Térmico do Cretáceo ocorreu no intervalo Cenomaniano-Turoniano, com auge por volta de 94 milhões de anos atrás. Foi o período mais quente dos últimos 145 milhões de anos: não havia gelo nos polos, florestas temperadas chegavam ao norte do Alasca e à Antártida, e a temperatura da superfície do mar nos trópicos e latitudes médias passava dos 35 °C, valor que hoje só é registrado no Golfo Pérsico no pico do verão.
A reconstrução de 485 milhões de anos de Judd, Tierney, Lunt e colaboradores (Science, 2024) confirma a escala: a temperatura média global da superfície (GMST) no pico do Cretáceo subiu para cerca de 35 a 36 °C, contra os 14 a 15 °C da média atual. Ou seja, o planeta inteiro era, em média, 20 °C mais quente do que é hoje. A curva longa de δ¹⁸O marinho do Fanerozoico mostra o mesmo sinal: o Cretáceo é um dos três grandes picos quentes da história da Terra, ao lado do Devoniano e do início do Triássico.
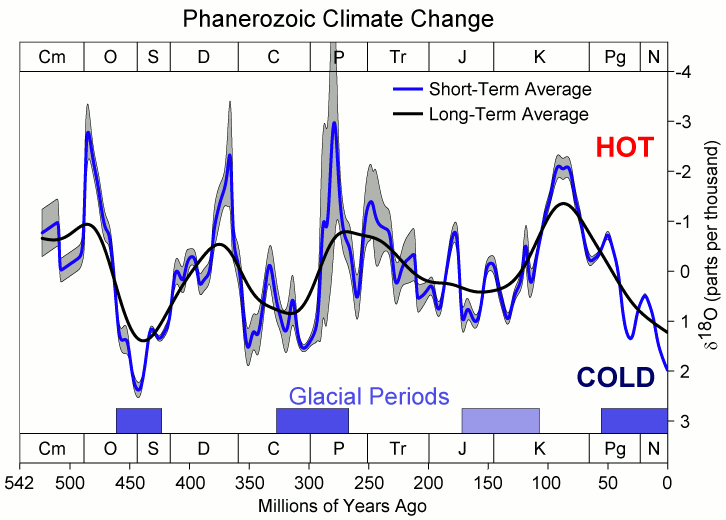
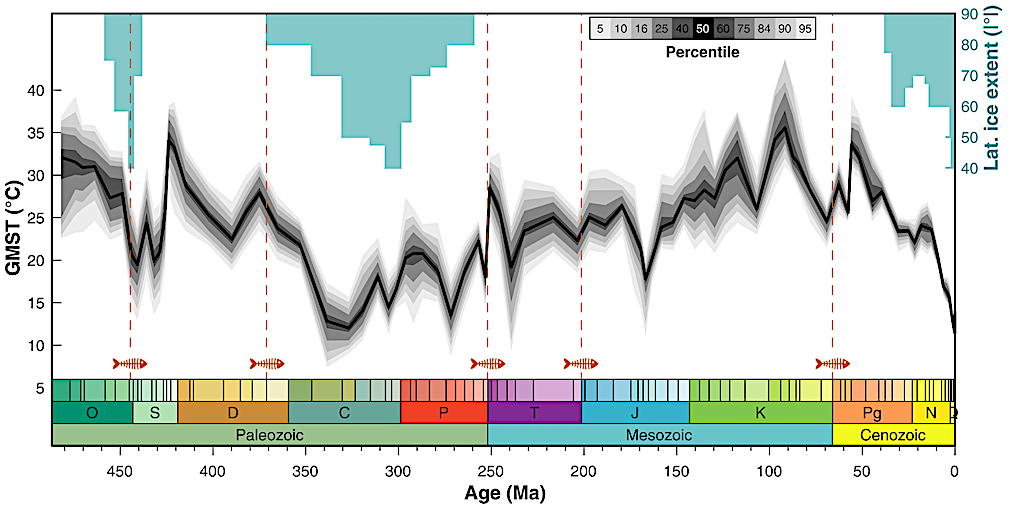
Depois desse pico, por volta de 90 a 85 milhões de anos atrás, as temperaturas globais começaram a cair lentamente. A queda continuou pelo Campaniano (83 a 72 Ma) e pelo Maastrichtiano (72 a 66 Ma), os últimos dois andares do Cretáceo, o mundo em que viveram o T. rex, o Tarbossauro e o Triceratops. Esses dinossauros terminais, portanto, habitaram um Cretáceo "já esfriando" em relação ao pico térmico, mas ainda com GMST cerca de 6 a 8 °C acima do atual, segundo a mesma reconstrução de Judd et al.
A pergunta então fica: onde estavam os maiores dinossauros? No pico do calor, ou depois, quando esfriou?
A resposta, levando a sério o registro fóssil, é direta: os maiores dinossauros conhecidos viveram justamente no pico do calor. O Argentinosaurus huinculensis, titanossauro da Patagônia, foi datado em 97 a 93 milhões de anos atrás, em plena zona do Máximo Térmico, e as estimativas de massa variam de 65 a 90 toneladas, provavelmente o maior animal terrestre já catalogado. O Patagotitan mayorum, outro titanossauro argentino, vem de camadas de cerca de 101 milhões de anos, ligeiramente antes do pico, com 55 a 77 toneladas. Entre os carnívoros do mesmo intervalo estão o Giganotosaurus, o Mapusaurus, o Carcharodontosaurus norte-africano e o Spinosaurus aegyptiacus, todos com 12 a 15 metros de comprimento.
Então o Cretáceo médio quente bate o Cretáceo terminal frio em gigantismo? Na linhagem dos saurópodes, sim. A coroa de "maior animal terrestre" pertence ao Cenomaniano. Um titanossauro de 90 toneladas é quase dez vezes mais pesado que um T. rex. Em termos puramente de massa, o pico de gigantismo do Reino dos Dinossauros foi no período mais quente, não no mais frio.
Mas isso não quer dizer que o calor tenha causado o gigantismo diretamente. A interpretação consensual é que o Máximo Térmico criou ecossistemas de produtividade ecológica extrema: florestas tropicais se estendendo por quase todo o planeta, mares epicontinentais rasos e quentes irrigando os continentes, plantas crescendo o ano inteiro, insolação forte o tempo todo. Essa biomassa vegetal gigantesca sustentou manadas de titanossauros de 60 a 90 toneladas, que por sua vez sustentaram predadores de 12 a 15 metros. Foi o clima que tornou possível um ecossistema desses, não o calor atuando nos corpos dos animais.
A prova disso está no que veio depois. Quando o planeta começou a esfriar ao final do Cenomaniano-Turoniano, os maiores titanossauros gondwânicos e os carcarodontossaurídeos se extinguiram. Alguns grupos encolheram, outros sumiram do registro. Mas o gigantismo não desapareceu: os tiranossaurinos (T. rex, Tarbossauro) e alguns titanossauros laurasianos como o Alamossauro preencheram os nichos abertos e atingiram tamanhos notáveis no Campaniano-Maastrichtiano, em clima mais frio. Só que em escala menor que os gigantes do pico térmico.
Resposta direta
Os maiores dinossauros de todos os tempos viveram no pico do calor (Cenomaniano-Turoniano, 100 a 90 milhões de anos atrás). Mas não foi o calor que os fez crescer: foi a produtividade ecológica que o clima quente e úmido tornou possível. Quando esfriou, alguns gigantes se extinguiram e outros apareceram (T. rex e companhia), mas nenhum chegou perto dos 70 a 90 toneladas dos titanossauros cenomanianos. Nem frio nem calor "fazem" o dinossauro crescer diretamente. O clima atua sempre pelos atalhos da produtividade de plantas, do tamanho das manadas de presas e da vaga ecológica deixada por quem se extingue.
Fontes: dados cronológicos e de massa de Argentinosaurus e Patagotitan compilados de Wikipedia/Argentinosaurus e Wikipedia/Patagotitan, baseados em Benson et al. 2014, Carballido et al. 2017 e Paul 2019. Temperaturas do Máximo Térmico do Cretáceo: Judd, Tierney, Lunt et al. 2024, Science 385, eadk3705, com dado complementar de Huber et al. 2018. Curva de δ¹⁸O do Fanerozoico adaptada de Veizer et al. 1999 e Prokoph et al. 2008. Extinção sincronizada de carcarodontossaurídeos e diplodocoides no Cenomaniano-Turoniano discutida em Novas et al. 2005 e Sales et al. 2016.
Quão grandes eram os dinossauros, exatamente?
Números em metros e toneladas são abstratos. Uma silhueta humana ao lado do animal transforma esses valores em escala imediata. O carrossel abaixo reúne 25 espécies do Jurássico e do Cretáceo, com comprimento e peso estimados pela literatura e as comparações visuais mais usadas em artigos científicos.
Sangue quente ou frio?
Durante décadas, paleontólogos debateram se dinossauros eram ectotermicos (sangue frio), como répteis, ou endotermicos (sangue quente), como aves e mamíferos. A resposta esta no meio, e o caminho até ela e uma das histórias mais fascinantes da paleontologia moderna.
O debate histórico
Até a década de 1960, o consenso era que dinossauros eram lagartos gigantes de sangue frio: lentos, dependentes do sol, limitados por temperaturas ambientais. Em 1968, Robert Bakker publicou um artigo que inaugurou a chamada "Renascença dos Dinossauros", argumentando que a postura ereta, as taxas de crescimento rápido e a proporção predador-presa nos ecossistemas mesozoicos eram incompatíveis com ectotermia. Bakker defendia que dinossauros eram tão endotérmicos quanto mamíferos. A visão tradicional, defendida por pesquisadores como R. Tracy e James Farlow, argumentava que a ectotermia explicaria a eficiência energética necessária para sustentar corpos tão grandes. A realidade, como quase sempre na biologia, se revelou mais complexa do que qualquer dos dois extremos.
O que os ossos revelam
A presença generalizada de osso fibrolamelar nos dinossauros indica crescimento rápido e continuo. Esse tipo de tecido ósseo se forma quando o animal cresce tão rápido que os osteocitos (celulas ósseas) ficam "presos" em uma matriz desordenada, sem tempo para se organizarem em camadas concentricas como no osso lamelar dos répteis. E o mesmo tipo de osso encontrado em aves e mamíferos. Ao mesmo tempo, muitas espécies apresentam Linhas de Crescimento Interrompido (LAGs, na sigla em ingles), pausas sazonais no crescimento que formam aneis visíveis em seções finas de osso, comparáveis aos aneis de árvores. Essas pausas são comuns em ectotermicos, que param de crescer durante estações frias ou secas. Essa combinação paradoxal, crescimento rápido com pausas sazonais, levou Grady et al. (2014) a proporem o conceito de mesotermia: um metabolismo intermediario entre répteis e mamíferos, após analisarem taxas de crescimento de 21 espécies de dinossauros comparadas a mais de 360 espécies vivas. Segundo o estudo, dinossauros tinham taxas metabolicas maiores que qualquer réptil vivo, mas menores que a maioria dos mamíferos.
Termometria isotopica
Estudos geoquimicos trouxeram evidências independentes. Eagle et al. (2011) utilizaram termometria de isotopos agrupados (clumped isotope thermometry) em cascas de ovos fossilizados. O principio e elegante: a frequência com que isotopos pesados de carbono-13 e oxigênio-18 se ligam no mesmo ion carbonato depende da temperatura em que o mineral se formou. Quanto mais fria a temperatura, maior a "preferência" desses isotopos pesados em se agruparem. Analisando cascas de ovos de saurópodes da Formação Allen (Patagonia, Argentina), Eagle estimou temperaturas corporais de 36 a 38°C, valores próximos aos de mamíferos modernos (37°C) e bem acima de répteis ectotermicos típicos (que acompanham a temperatura ambiente). Dawson et al. (2020) expandiram essa abordagem, analisando isotopos de oxigênio em cascas de ovos de ambas as linhagens de dinossauros (Ornithischia e Saurischia), concluindo que temperaturas corporais elevadas em relação ao ambiente eram provavelmente uma característica ancestral do grupo inteiro, presente desde antes da diversificação dos dinossauros.
Variação entre grupos
A fisiologia termica variava enormemente dentro de Dinosauria. Teropodes pequenos, ancestrais das aves, eram provavelmente endotermicos plenos, com isolamento termico fornecido por penas e taxas metabolicas altas o suficiente para manter temperatura corporal constante independentemente do ambiente. Grandes saurópodes podem ter mantido temperaturas estáveis por um mecanismo completamente diferente: gigantotermia (homeotermia inercial). A massa corporal enorme funciona como um reservatorio termico: um animal de 30 toneladas tem uma relação superficie-volume tão baixa que leva dias para perder calor significativamente, mesmo sem gerar calor metabolico ativo. Esse fenômeno e observado hoje na tartaruga-de-couro (Dermochelys coriacea), que mantem temperatura central 8-10°C acima da água ao redor, apesar de ser ectotermia. Para dinossauros medios, como ornitopodes de 500 kg a 2 toneladas, a situação era mais ambigua: provavelmente geravam calor metabolico ativo, mas com menos eficiência que mamíferos. A divisão simplificada entre "sangue quente" e "sangue frio" e inadequada para descrever a diversidade fisiológica desses animais.
Turbinatos nasais: a pista que não se preservou
Mamíferos e aves possuem estruturas ósseas finas dentro da cavidade nasal chamadas turbinatos respiratorios. Essas estruturas aquecem e umidificam o ar inspirado e recuperam água do ar expirado, essenciais para animais endotermicos que perdem muita água pela respiração rápida. Ruben et al. (1996) argumentaram que a ausência de turbinatos em dinossauros indicava ectotermia. O problema e que essas estruturas são extremamente frágeis e raramente se preservam em fósseis, mesmo em mamíferos. Tomografias computadorizadas de crânios de dinossauros revelam cavidades nasais complexas com cristas e sulcos que poderiam ter sustentado tecido mole equivalente, mas a evidência permanece inconclusiva. A ausência de evidência, nesse caso, não e evidência de ausência.



Penas ou escamas?
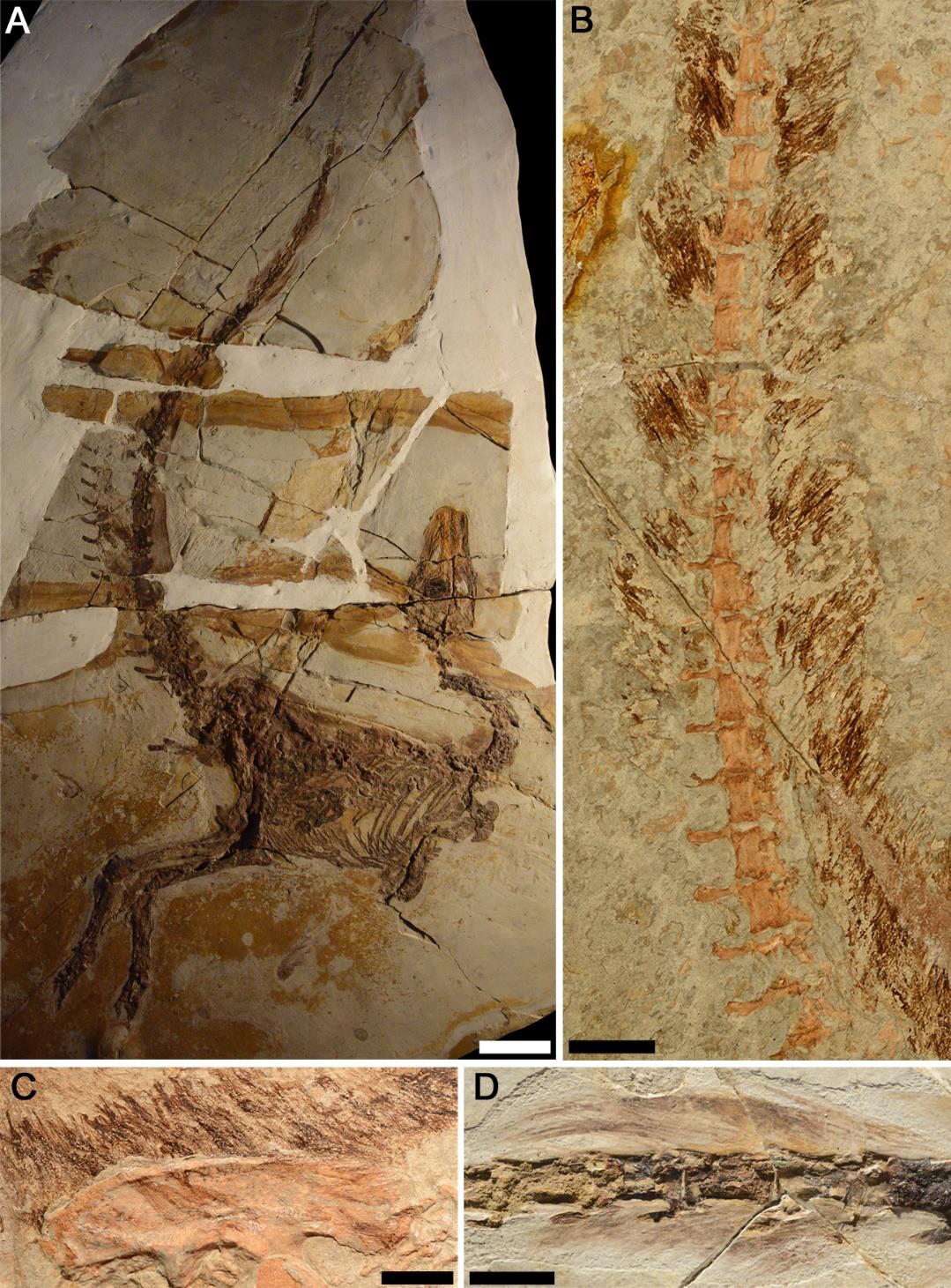
A descoberta de dinossauros emplumados em Liaoning, China, a partir da década de 1990, revolucionou a paleontologia. Penas não eram exclusivas das aves, e suas funções originais nada tinham a ver com voar.
A revolução de Liaoning
Em 1996, o Sinosauropteryx foi descrito como o primeiro dinossauro não aviano com estruturas tegumentares semelhantes a penas (proto-penas). O fóssil, preservado em calcário de grão finíssimo da Formação Yixian, mostrava claramente um halo de filamentos ao redor do corpo, incluindo padrões de contra-sombreamento (dorso escuro, ventre claro) que indicam coloração funcional. Desde então, dezenas de espécies emplumadas foram encontradas nos depositos vulcanicos de Liaoning, que preservam tecidos moles com detalhe extraordinário graças ao soterramento rápido por cinzas vulcanicas em lagos rasos. Esse tipo de deposito geológico e chamado de Lagerstatte (termo alemão para "local de armazenamento"), e Liaoning e considerado o mais importante do mundo para dinossauros emplumados.
Espécies-chave
O Microraptor gui (2003) e um dromeossaurideo com quatro "asas", penas de voo asimetricas tanto nos membros anteriores quanto nos posteriores, que evidenciam capacidade de planar ou até voo rudimentar. Analises aerodinamicas sugerem que ele planava entre árvores, usando as penas traseiras como estabilizadores. O Anchiornis huxleyi (2009) preservou melanossomas (organelas que contem pigmentos) tão bem que pesquisadores reconstruiram sua coloração completa: corpo cinza-escuro, crista vermelha, penas de voo brancas com pontas pretas. Foi o primeiro dinossauro com coloração mapeada corpo inteiro. O Yutyrannus huali (2012) e o maior dinossauro emplumado conhecido: um tiranossauroide de ~9 metros e ~1,4 toneladas com filamentos cobrindo grande parte do corpo. Sua existência demonstrou que penas não eram exclusivas de dinossauros pequenos.
Além dos terópodes
Em 2014, o Kulindadromeus zabaikalicus, encontrado na Russia, ampliou o cenario: trata-se de um ornitisquio com estruturas semelhantes a penas, incluindo três tipos distintos de coberturas filamentosas. Ornitisquios são a linhagem oposta aos terópodes na árvore filogenetica dos dinossauros. A presença de estruturas plumosas em ambas as linhagens sugere que alguma forma de cobertura filamentosa pode ser ancestral a todos os dinossauros, surgindo uma única vez no ancestral comum do grupo inteiro. Se isso for correto, dinossauros escamosos como saurópodes e ceratopsianos teriam perdido as estruturas filamentosas secundariamente, não o contrario. A ideia ainda e debatida, mas ganhou força após a descrição de Kulindadromeus.
Estagio evolutivo das penas (modelo de Prum)
Richard Prum (1999) propôs um modelo de cinco estagios para a evolução das penas, do mais simples ao mais complexo: I. Filamento único oco (como em Sinosauropteryx). II. Filamentos ramificados a partir de uma base (plumulaceas). III. Barbulas com ganchos que travam as barbas entre si (penas planares). IV. Assimetria entre as barbas (funcional para aerodinamica). V. Diversificação adicional (penas especializadas de voo, exibição, etc.). Esse modelo predizia a existência de penas em estagios intermediarios anos antes de fósseis como Anchiornis e Kulindadromeus confirmarem estruturas nos estagios I e II em dinossauros não avianos.
O caso do T. rex: penas ou escamas?
O próprio T. rex é alvo de debate intenso. Impressões de pele publicadas por Bell et al. (2017) revelaram escamas em áreas como abdômen, cauda e pescoço, com texturas similares à pele de crocodilos. Isso indicaria que tiranossauros de grande porte podem ter perdido penas secundariamente, possivelmente por pressão termorreguladora: animais grandes em climas quentes precisam dissipar calor, e penas atrapalham. É o mesmo princípio que explica por que elefantes e rinocerontes perderam pelos. No entanto, as impressões de pele de Bell cobrem apenas ~1% da superfície corporal, o que deixa aberta a possibilidade de que T. rex tivesse penas em regiões não preservadas, como o dorso ou a crista.
Funções originais das penas
As penas não evoluiram para o voo. Evidencias apontam para multiplas funções anteriores: exibição sexual e comunicação visual (padrões de cor preservados em fósseis, incluindo iridescencia em Microraptor, detectada por Li et al. em 2012); isolamento termico (particularmente importante para dinossauros pequenos endotermicos em climas frios); proteção de ovos durante o choco (oviraptorídeos fossilizados sobre ninhos com braços abertos, em posição identica a aves modernas que cobrem ovos com penas); e possivelmente camuflagem (contra-sombreamento em Sinosauropteryx). O voo surgiu depois, como uma cooptação funcional de estruturas já existentes, provavelmente passando por estagios intermediarios de voo planado, como visto em Microraptor.
Distribuição por grupo
Com penas
Celurossauros (tiranossauroides, dromeossaurideos, aves)
Com escamas
Sauropodes, anquilossauros, ceratopsianos (maioria)


Solitarios ou em bando?
Os dinossauros exibiam ampla gama de comportamentos sociais: desde habitos solitários até manadas com milhares de individuos. Decifrar o comportamento de animais extintos e um dos maiores desafios da paleontologia, mas o registro fóssil fornece pistas surpreendentemente detalhadas.
Bonebeds: cemitérios coletivos
As evidências mais robustas de vida gregaria vem dos bonebeds (jazigos ósseos): depositos onde centenas ou milhares de individuos da mesma espécie foram preservados juntos. O Hilda bonebed em Alberta, Canada, contem restos de mais de 1.000 individuos de Centrosaurus apertus espalhados por 2,3 km², interpretado como uma manada inteira morta durante uma enchente ao tentar cruzar um rio (Eberth & Getty, 2005). Os bonebeds de Maiasaura na Formação Two Medicine, Montana, EUA, com até 10.000 individuos estimados, indicam manadas enormes que provavelmente migravam sazonalmente (Varricchio & Horner, 1993). A questão critica na interpretação de bonebeds e a tafonomia: como distinguir entre animais que viviam juntos e morreram simultaneamente versus cadaveres transportados por água e acumulados no mesmo local ao longo do tempo? A presença de individuos de todas as faixas etarias (filhotes a adultos) e a ausência de segregação por tamanho no deposito favorecem a interpretação de morte em massa de um grupo social coeso.
Trilhas e migração
Trilhas fossilizadas paralelas de saurópodes, como as do rio Purgatoire no Colorado (mais de 1.300 pegadas em 100 trilhas), confirmam que grupos caminhavam juntos na mesma direção e na mesma velocidade. A análise dessas trilhas permite calcular velocidades individuais e verificar se os animais se moviam de forma coordenada. No sitio de Cal Orcko, Bolivia, mais de 5.000 pegadas de 462 trilhas de multiplas espécies cobrem uma parede calcaria de 25.000 m², um dos maiores registros de atividade de dinossauros do mundo. Trilhas de saurópodes em sitios como Davenport Ranch, Texas, mostram pegadas de individuos pequenos no centro e grandes nas bordas, sugerindo comportamento protetor semelhante ao de manadas de elefantes, que posicionam filhotes no meio do grupo durante deslocamentos.
Cuidado parental
O cuidado parental também é bem documentado. Maiasaura peeblesorum, cujo nome significa "lagarto boa mãe", foi descrita por Jack Horner em 1979 a partir de colônias de ninhos em Egg Mountain, Montana. Os ninhos estavam espaçados cerca de 7 metros (aproximadamente o comprimento do animal adulto), com filhotes que permaneciam no ninho até dobrar de tamanho, evidenciado pelo desgaste dental dos filhotes (eles já estavam comendo, mas ainda não caminhavam). Oviraptorídeos como Citipati osmolskae foram encontrados fossilizados diretamente sobre seus ninhos em posição de incubação, braços estendidos sobre os ovos como aves modernas. Em 2021, Pol et al. publicaram a descoberta de uma colônia de nidificação de Mussaurus patagonicus na Patagônia argentina, com 80 ovos e 100 esqueletos organizados por faixa etária: adultos em um setor, juvenis em outro, ovos em um terceiro. Isso sugere segregação etária dentro de um grupo social, algo inédito no registro fóssil de dinossauros.
Comunicação
Vida social requer comunicação, e os dinossauros tinham estruturas anatomicas sofisticadas para isso. A crista oca de Parasaurolophus funcionava como câmara de ressonancia, produzindo sons de baixa frequência (~30-700 Hz) para comunicação a distância (Weishampel, 1981). Reconstruções digitais do tubo interno da crista permitiram simular o som provável: um mugido grave, similar ao de um instrumento de sopro. Escudos e chifres de ceratopsianos como Triceratops provavelmente serviam para reconhecimento visual de espécie e seleção sexual (Padian & Horner, 2011). A enorme variação na forma dos escudos entre espécies próximas (ex: Triceratops vs. Torosaurus vs. Chasmosaurus) apoia essa hipótese. Em hadrossauros, a diversidade de formatos de crista dentro da mesma família sugere que cada espécie tinha um "canal sonoro" próprio, permitindo individuos de espécies diferentes coexistir sem confusão de sinais.
Teropodes: solitários ou gregários?
A maioria dos terópodes grandes, como T. rex e Allosaurus, eram provavelmente solitários ou territoriais, semelhantes a grandes felinos modernos. No entanto, existem exceções. Sítios com multiplos espécimes de Mapusaurus na Argentina e de Albertosaurus em Alberta sugerem algum grau de agregação social em terópodes grandes. Currie (1998) descreveu um bonebed com 12 individuos de Albertosaurus sarcophagus de diferentes idades, que ele interpretou como um grupo social. A interpretação e debatida: os animais podem ter sido atraidos independentemente a uma carcaça ou fonte de água, como ocorre com crocodilos modernos, sem que houvesse cooperação real.


Como caçavam os predadores?
Maquinas de caça refinadas por 165 milhões de anos de evolução. Cada grupo de predadores desenvolveu estrategias distintas, da mordida esmagadora do T. rex a precisão cirurgica dos raptores e a caça aquatica dos espinossaurideos.
A mordida do T. rex
O T. rex possuía a mordida mais poderosa já registrada em um animal terrestre: entre 35.000 e 57.000 newtons, o equivalente a quase 6 toneladas de força concentradas em um único dente posterior (Bates & Falkingham, 2012). Para comparação, um leão moderno gera ~4.000 N e um crocodilo-de-água-salgada ~16.000 N. Essa pressão era suficiente para esmagar ossos e extrair a medula, um comportamento chamado osteovagia, confirmado por coprolitos (fezes fossilizadas) de T. rex contendo fragmentos ósseos triturados (Chin et al., 1998). Seus dentes serrilhados (dentição zifodonte) funcionavam como facas de churrasco: as micro-serrilhas reduziam a força necessária para cortar carne, enquanto estruturas internas de dentina reforçavam cada denticulo contra fraturas. Cada dente tinha cerca de 15-30 cm de comprimento, com raizes profundas ancoradas no maxilar por ligamentos periodontais robustos. Os dentes eram substituidos continuamente: quando um caia, outro já estava crescendo por baixo, um ciclo que durava cerca de 2 anos por dente.
Sentidos e locomoção
A visão binocular do T. rex cobria cerca de 55 graus (Stevens, 2006), com olhos de 13 cm de diâmetro, maiores que os de qualquer animal terrestre vivo. Tomografias computadorizadas do crânio revelam bulbos olfatorios proporcionalmente enormes, sugerindo um olfato comparável ao de abutres modernos, capazes de detectar cadaveres a quilômetros de distância. O ouvido interno indica sensibilidade a sons de baixa frequência, úteis para localizar presas grandes. A prova definitiva de predação ativa, e não apenas consumo de carcaças, veio em 2013: um dente de T. rex foi encontrado cravado entre vértebras cicatrizadas de um hadrossauro (Edmontosaurus), demonstrando que a presa estava viva quando foi atacada e sobreviveu ao ataque (DePalma et al., 2013). Apesar do porte, T. rex provavelmente não era rápido: estimativas biomecanicas indicam 20-29 km/h como velocidade máxima sustentavel (Hutchinson & Garcia, 2002). Acima disso, os ossos das pernas sofreriam fraturas por estresse. Suas presas, como Edmontosaurus, eram igualmente lentas, o que tornava a velocidade relativa adequada.
Estrategias alternativas: Allosaurus e Spinosaurus
Nem todos os predadores funcionavam como T. rex. O Allosaurus, predador dominante do Jurássico, tinha uma mandíbula mais fraca, mas um crânio leve e músculos cervicais potentes. Rayfield et al. (2001) propuseram a hipótese do "golpe de machadinha": Allosaurus abria a boca em angulo de ~80 graus e descia o crânio contra a presa com força, usando o maxilar superior como arma cortante. Os dentes curvos e serrilhados rasgavam a carne no impacto, causando hemorragia massiva. Spinosaurus aegyptiacus, por sua vez, era um predador semiaquatico. Ibrahim et al. (2014, 2020) demonstraram que seu focinho longo e conico, repleto de receptores de pressão semelhantes aos de crocodilos, e sua cauda achatada lateralmente (como a de uma salamandra gigante) indicam adaptação para caça de peixes e outros animais aquaticos. Os dentes conicos e sem serrilhas eram ideais para prender presas escorregadias, não para cortar carne.
Modelo RPR: como os raptores caçavam
Fowler et al. (2011) propuseram o modelo Raptor Prey Restraint (RPR): dromeossaurideos como o Deinonychus saltavam sobre a presa, prendiam-na com as garras curvas do segundo dedo (a famosa "garra retratil") e a consumiam ainda viva, batendo os braços emplumados para manter o equilibrio enquanto a presa tentava escapar. Exatamente como aguias e gavioes fazem hoje ao capturar presas maiores que eles. A garra do segundo dedo, que popularmente se acredita ser uma arma cortante, na verdade funcionava mais como um gancho de fixação: análises biomecanicas mostram que a ponta era otimizada para perfurar e prender, não para rasgar (Manning et al., 2009). O dano principal vinha das mandíbulas, que se alimentavam da presa imobilizada.
Caça em matilha: mito?
A ideia de que dromeossaurideos caçavam em matilhas coordenadas como lobos, popularizada pelo filme Jurassic Park (1993), foi contestada por Roach & Brinkman (2007). Analisando sitios como a Pedreira Antlers em Oklahoma, onde multiplos espécimes de Deinonychus foram encontrados junto a um Tenontosaurus, eles encontraram evidências de canibalismo: dentes de Deinonychus cravados em ossos de outros Deinonychus. Isso sugere um comportamento mais parecido com dragoes de Komodo: alimentação oportunista em grupo, com agressão entre individuos, sem cooperação real. Dragoes de Komodo se congregam ao redor de grandes carcaças e frequentemente se atacam mutuamente. Estudos de isotopos dentarios e padrões de desgaste dentario em sitios multi-individuos de terópodes apoiam essa interpretação (Frederickson et al., 2020).
O debate caçador vs. carniceiro
Jack Horner defendeu durante anos que T. rex era primariamente um carniceiro, baseando-se em seus braços atrofiados, olfato potente (útil para encontrar carcaças) e velocidade relativamente baixa. A maioria dos paleontólogos hoje rejeita essa hipótese como exclusiva: nenhum grande predador terrestre conhecido e carniceiro obrigatorio (nem abutres são exclusivamente carniceiros). O consenso atual e que T. rex era um predador oportunista, como leoes e hienas, que caçava quando possível e se alimentava de carcaças quando disponíveis. O dente cicatrizado no hadrossauro e a evidência mais forte de predação ativa.



Quanto tempo viviam?
Paleontologistas determinam a idade dos dinossauros pela histologia óssea: cortam ossos longos em seções finas (30-100 micrometros) e contam os aneis de crescimento sob microscopio, como em árvores. Cada anel representa aproximadamente um ano de vida.
Como funciona a histologia óssea
O método consiste em extrair uma seção transversal de um osso longo (geralmente a tíbia ou o fêmur), embuti-la em resina, corta-la em lâminas de ~30 micrometros de espessura com um microtomo, e analisa-la sob luz polarizada. As Linhas de Crescimento Interrompido (LAGs) aparecem como aneis escuros e densos, formados durante períodos de crescimento reduzido (inverno, estação seca). Entre as LAGs, o tecido ósseo pode ser fibrolamelar (crescimento rápido) ou lamelar (crescimento lento). A espessura entre LAGs consecutivas diminui conforme o animal envelhece, formando uma curva de crescimento sigmoidal: crescimento lento na infancia, aceleração explosiva na adolescencia, desaceleração na maturidade, e estagnação na velhice. Um dos problemas do método e que a remodelação óssea (reabsorção e redeposição de tecido ósseo pelo corpo) apaga as LAGs internas ao longo da vida, subestimando a idade de animais velhos. Sauropodes são particularmente afetados por isso.
Curvas de crescimento de terópodes
O espécime "Sue" de T. rex morreu com aproximadamente 28 anos e atingiu o tamanho adulto por volta dos 19. No pico de crescimento, entre 14 e 18 anos, ganhava cerca de 2,1 kg por dia (Erickson et al., 2004), o equivalente a passar de ~500 kg para mais de 5.000 kg em apenas 4 anos. Esse crescimento explosivo e uma das evidências mais fortes de metabolismo elevado. Em comparação, Albertosaurus atingia o tamanho adulto com ~16 anos e vivia até ~28, enquanto Gorgosaurus crescia mais rápido, mas morria mais jovem. A vida curta de tiranossaurideos sugere que eram animais de alto metabolismo e alto risco: cresciam rápido, reproduziam cedo, e morriam relativamente jovens. O Psittacosaurus, um ceratopsiano basal de 2 metros, e o dinossauro não aviano com mais dados histologicos disponíveis. Erickson et al. (2009) analisaram dezenas de espécimes e determinaram que atingia a maturidade com 8-9 anos e vivia até ~10-11 anos.
Reprodução precoce
A maturidade sexual vinha antes da esquelética. Osso medular, um tecido reprodutivo rico em calcio que hoje só existe em aves femeas durante o período de postura (fornece calcio para as cascas de ovos), foi encontrado em T. rex, Allosaurus e Tenontosaurus, todos em espécimes ainda em crescimento ativo (Lee & Werning, 2008). Isso significa que dinossauros já se reproduziam antes de parar de crescer: um padrão mais próximo de aves do que de répteis, que tipicamente atingem maturidade sexual apenas após o crescimento completo. A reprodução precoce e uma estrategia classica de animais com alta mortalidade: começar a reproduzir cedo aumenta as chances de deixar descendentes antes de morrer.
Mortalidade juvenil e tabela de vida
O estudo de Woodward et al. (2015) com 50 tíbias de Maiasaura e o exemplo mais completo de paleodemografia em dinossauros. Ao determinar a idade de cada individuo pela histologia, os pesquisadores construiram uma tabela de vida completa, algo que normalmente só e possível para populações vivas. Os resultados revelaram três fases distintas: mortalidade de 89,9% no primeiro ano de vida, provavelmente por predação, doença e inanição; um período de 7 anos de "janela de sobrevivencia" com mortalidade relativamente baixa (12,7% ao ano), quando os animais já eram grandes o suficiente para evitar a maioria dos predadores; e senescencia após os 8 anos, quando a taxa subia para 44,4%, provavelmente por acumulo de lesoes, doenças e desgaste fisiológico. Maiasaura chegava a 2,3 toneladas em apenas 8 anos.
Patologias: o que os ossos doentes revelam
Os ossos dos dinossauros frequentemente preservam sinais de doenças e lesoes. "Sue" sofria de gota, infecções ósseas (osteomielite) e possíveis mordidas infectadas de outros T. rex. Fraturas cicatrizadas em costelas e vértebras são comuns em terópodes grandes, indicando que sobreviviam a lesoes que seriam fatais para animais menores. Em Allosaurus, mais de 50% dos espécimes do Cleveland-Lloyd Quarry apresentam fraturas cicatrizadas. Essas patologias fornecem informações sobre longevidade (o animal viveu tempo suficiente para curar), comportamento (brigas intraespecificas) e mesmo dieta (gota esta associada a dietas ricas em purinas, presentes em carne).
~28
anos
T. rex
~8
anos até adulto
Maiasaura
30-70
anos (estimado)
Sauropodes


Quão inteligentes eram?
Medir a inteligência de animais extintos e um dos maiores desafios da paleontologia. O cerebro, feito de tecido mole, não se preserva em fósseis. O que temos são endocasts: moldes naturais ou digitais (via tomografia computadorizada) da cavidade craniana, que revelam o tamanho e a forma geral do cerebro.
Quociente de Encefalização (QE)
O QE, proposto por Harry Jerison em 1973, mede a proporção entre a massa do encefalo e o esperado para um animal daquele tamanho corporal. A lógica e que animais maiores precisam de cerebros maiores apenas para controlar seus corpos: o que "sobra" além desse mínimo pode ser correlacionado com capacidades cognitivas. Entre os dinossauros não aviarios, o Troodon formosus detém o maior QE registrado: seu encefalo era cerca de seis vezes maior do que o previsto para um réptil de mesmo porte, valor comparável ao de emas e avestruzes modernas. Troodon tinha olhos grandes voltados para frente (visão binocular), garras retrateis nos pes, e mãos preensiveis com oposição parcial do polegar. No extremo oposto, saurópodes como Brachiosaurus carregavam encefalo de apenas 70 a 100 cm³ em corpos de mais de 30 toneladas (para comparação, um gato domestico tem ~25 cm³ em um corpo de 4 kg). O QE tem limitações sérias: ele assume que a relação cerebro-corpo segue uma única formula para todos os animais, o que não e verdade. Aves, por exemplo, empacotam mais neuronios por grama de cerebro do que mamíferos.
O cerebro do T. rex
O T. rex possuía um encefalo surpreendentemente grande (~400 cm³), com bulbos olfatorios e hemisferios cerebrais bem desenvolvidos. Os lobos olfatorios ocupavam cerca de 40% do volume cerebral, indicando um olfato excepcional. O cerebelo, responsável pela coordenação motora, também era proporcionalmente grande, consistente com um predador que precisava de movimentos precisos de cabeça e corpo durante o ataque. Os lobos opticos eram moderados, sugerindo que a visão, embora boa, não era o sentido dominante.
A controversia Herculano-Houzel
Em 2023, a neurocientista brasileira Suzana Herculano-Houzel publicou um estudo estimando que T. rex poderia ter tido ~3,3 bilhões de neuronios no telencefalo, número comparável ao de babuínos e macacos-prego, animais capazes de usar ferramentas e resolver problemas complexos. Se correto, isso implicaria que T. rex era cognitivamente sofisticado, possivelmente capaz de comportamentos que antes eram considerados exclusivos de primatas. A metodologia, porém, foi severamente criticada por Gutierrez-Ibañez et al. (2023) e por Caspar et al. (2024). As criticas principais: Herculano-Houzel usou equações de escalonamento neuronal derivadas de aves para estimar neuronios em répteis. Aves empacotas neuronios de forma muito mais densa que répteis. Usar a equação de aves superestima drasticamente a contagem de neuronios. Os criticos recalcularam usando equações de répteis e chegaram a ~1,5 bilhão de neuronios, número comparável a crocodilos grandes, não a primatas. O tamanho do encefalo usado por Herculano-Houzel também foi questionado: ela usou o volume total do endocast, mas em répteis o cerebro ocupa apenas 30-50% da cavidade craniana (o restante e preenchido por meninges e fluido). Esse resultado permanece altamente controverso e a maioria dos paleontólogos se posiciona com cautela.
Mito: o "segundo cerebro"
No século XIX, O.C. Marsh observou que o canal neural na região sacral de Stegosaurus era 20 vezes maior que a cavidade craniana. A imprensa da época popularizou a ideia de um "segundo cerebro no quadril". Na realidade, esse alargamento não era um segundo cerebro. Estudos comparativos com aves modernas (Giffin, 1991) mostraram que o alargamento sacral em aves e ocupado pelo corpo glicogenico, uma estrutura rica em glicogenio (reserva de energia) cuja função exata ainda e debatida, mas que esta ligada ao metabolismo energético do sistema nervoso, não ao processamento neural. Sauropodes e ornitisquios provavelmente tinham estruturas analogas. O "segundo cerebro" e, portanto, um mito vitoriano que persiste na cultura popular.
Capacidades sensoriais
Endocasts e tomografias computadorizadas revelam informações sobre os sentidos dos dinossauros. O ouvido interno preservado em crânios permite estimar a faixa de frequência auditiva: Parasaurolophus era sensível a sons de baixa frequência (consistente com as vocalizações da crista), enquanto terópodes pequenos como Troodon ouviam frequências mais altas, úteis para localizar presas pequenas. A flocula, uma estrutura do cerebelo visível em endocasts, e proporcional a agilidade do olhar: animais com floculas grandes podem manter o olhar estável enquanto movem a cabeça rapidamente. Teropodes manirraptores (dromeossaurideos, troodontideos) tinham floculas proporcionalmente maiores que qualquer outro dinossauro, sugerindo agilidade visual comparável a aves de rapina.
Comportamento complexo
Não há evidência confirmada de uso de ferramentas em nenhum dinossauro não aviano. Resolução de problemas e aprendizagem social permanecem especulativos, inferidos apenas indiretamente a partir de acumulações ósseas que sugerem comportamento gregário e cuidado parental. O que sabemos e que os terópodes manirraptores, especialmente troodontideos, tinham encefalo, sentidos e destreza manual compatíveis com comportamentos relativamente sofisticados, possivelmente comparáveis aos de corvos ou papagaios modernos. No entanto, extrapolar cognição a partir de anatomia e inerentemente incerto, e afirmações sobre dinossauros "tão inteligentes quanto primatas" devem ser tratadas com ceticismo até que evidências mais robustas surjam.



Referencias
Bates, K.T. & Falkingham, P.L. (2012). Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics. Biology Letters, 8(4), 660-664.
Bell, P.R. et al. (2017). Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution. Biology Letters, 13(6), 20170092.
Caspar, K.R. et al. (2024). How smart was T. rex? Testing claims of exceptional cognition in dinosaurs. The Anatomical Record, 307(4), 1223-1241.
Chen, P. et al. (1998). An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature, 391, 147-152.
Chin, K. et al. (1998). A king-sized theropod coprolite. Nature, 393, 680-682.
Dawson, R.R. et al. (2020). Eggshell geochemistry reveals ancestral metabolic thermoregulation in Dinosauria. Science Advances, 6(7), eaax9361.
Eagle, R.A. et al. (2011). Body temperatures of modern and extinct vertebrates from ¹³C-¹⁸O bond abundances in bioapatite. PNAS, 108(25), 10255-10260.
Eberth, D.A. & Getty, M.A. (2005). Ceratopsian bonebeds: occurrence, origins, and significance. In Currie, P.J. & Koppelhus, E.B. (eds.), Dinosaur Provincial Park, Indiana Univ. Press.
Erickson, G.M. et al. (2004). Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature, 430, 772-775.
Erickson, G.M. et al. (2009). A life table for Psittacosaurus lujiatunensis. Proc. Royal Society B, 276, 3085-3090.
Fowler, D.W. et al. (2011). The predatory ecology of Deinonychus and the origin of flapping in birds. PLOS ONE, 6(12), e28964.
Frederickson, J.A. et al. (2020). Ontogenetic dietary shifts in Deinonychus antirrhopus. Palaeogeography, Palaeoclimatology, Palaeoecology, 552, 109780.
Giffin, E.B. (1991). Endosacral enlargements in dinosaurs. Modern Geology, 16, 101-112.
Grady, J.M. et al. (2014). Evidence for mesothermy in dinosaurs. Science, 344(6189), 1268-1272.
Gutierrez-Ibañez, C. et al. (2023). How smart was T. rex? The Anatomical Record.
Horner, J.R. & Makela, R. (1979). Nest of juveniles provides evidence of family structure among dinosaurs. Nature, 282, 296-298.
Hutchinson, J.R. & Garcia, M. (2002). Tyrannosaurus was not a fast runner. Nature, 415, 1018-1021.
Ibrahim, N. et al. (2014, 2020). Semiaquatic adaptations in a giant predatory dinosaur. Science, 345, 1613-1616; Nature, 581, 67-70.
Jerison, H.J. (1973). Evolution of the Brain and Intelligence. Academic Press.
Lee, A.H. & Werning, S. (2008). Sexual maturity in growing dinosaurs does not fit reptilian growth models. PNAS, 105(2), 582-587.
Li, Q. et al. (2012). Reconstruction of Microraptor and the evolution of iridescent plumage. Science, 335(6073), 1215-1219.
Manning, P.L. et al. (2009). Biomechanics of dromaeosaurid dinosaur claws. Proc. Royal Society B, 276, 1071-1079.
Pol, D. et al. (2021). Earliest evidence of herd-living and age segregation amongst dinosaurs. Scientific Reports, 11, 20023.
Prum, R.O. (1999). Development and evolutionary origin of feathers. J. Experimental Zoology, 285(4), 291-306.
Rayfield, E.J. et al. (2001). Cranial design and function in a large theropod dinosaur. Nature, 409, 1033-1037.
Roach, B.T. & Brinkman, D.L. (2007). A reevaluation of cooperative pack hunting in Deinonychus. Bulletin Peabody Museum, 48(1), 103-138.
Ruben, J.A. et al. (1996). The metabolic status of some Late Cretaceous dinosaurs. Science, 273, 1204-1207.
Stevens, K.A. (2006). Binocular vision in theropod dinosaurs. J. Vertebrate Paleontology, 26(2), 321-330.
Varricchio, D.J. & Horner, J.R. (1993). Hadrosaurid and lambeosaurid bone beds from the Two Medicine Formation. Canadian Journal of Earth Sciences, 30(5), 997-1006.
Woodward, H.N. et al. (2015). Maiasaura, a model organism for extinct vertebrate population biology. Paleobiology, 41(4), 503-527.
Xu, X. et al. (2012). A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature, 484, 92-95.