Dakotaraptor
Dakotaraptor steini
"Ladrão de Dakota, de Stein"
Sobre esta espécie
Dakotaraptor steini é um grande dromeossaurídeo do Cretáceo Superior (Maastrichtiano tardio, cerca de 66 a 67 milhões de anos atrás) da Formação Hell Creek, em Harding County, South Dakota. Descrito por Robert DePalma, David Burnham, Larry Martin, Peter Larson e Robert Bakker em 2015, o animal atingia entre 4,35 e 6 metros de comprimento, com massa corporal estimada entre 220 e 350 quilogramas, sendo o maior dromeossaurídeo conhecido do Maastrichtiano norte-americano e um dos maiores da família, ao lado de Utahraptor e Achillobator. O holótipo PBMNH.P.10.113.T consiste em um esqueleto parcial de um adulto sem crânio, descoberto por Robert DePalma em 2005 em um canal fluvial do Hell Creek, a no máximo 20 metros abaixo do limite Cretáceo-Paleógeno. A ulna preserva cerca de 15 papilas ulnares (quill knobs) de 8 a 10 milímetros de diâmetro, indicando inserção de penas grandes, e as garras do segundo dedo do pé, a clássica 'sickle claw' dos dromeossaurídeos, alcançam 24 centímetros de comprimento medido pela curva externa. A tíbia, com 678 milímetros, é a mais longa conhecida em qualquer dromeossaurídeo, sugerindo anatomia surpreendentemente esguia para um animal desse porte. Em 2016, Arbour e colegas demonstraram que as fúrculas originalmente descritas como parte do holótipo eram, na verdade, entoplastros (partes do casco) da tartaruga trionichídea Axestemys splendida, e DePalma et al. (2016) publicaram um corrigendum excluindo essas peças. A validade do táxon segue debatida: Cau (2023 e 2024) argumentou, em blog e análises filogenéticas, que o material restante pode ser uma quimera combinando elementos de ornitomimossauros, ovirraptorossauros e terizinossauros, mas essa hipótese ainda não foi publicada em artigo revisado por pares.
Formação geológica e ambiente
Formação Hell Creek, topo da unidade, Cretáceo Superior (Maastrichtiano tardio, ~66 a 67 Ma). A Formação Hell Creek aflora em Montana, North Dakota, South Dakota e Wyoming, registrando sedimentos fluviais e de planície costeira depositados nos últimos 1 a 2 milhões de anos do Cretáceo, imediatamente antes do limite Cretáceo-Paleógeno (K-Pg), datado em 66,043 ± 0,010 Ma por Sprain et al. (2018). O litotipo inclui arenitos de canal, mudstones de planície de inundação, carvões de pântano e bentonitas vulcanoclásticas. A unidade ocorre sobre a Formação Fox Hills (marinha) e abaixo da Formação Tullock / Fort Union (Paleoceno). Em Harding County, South Dakota, a Formação Hell Creek inclui o arenito Bull Creek e o carvão Heikkila, e é nesse intervalo que o holótipo de Dakotaraptor foi encontrado, a menos de 20 metros abaixo do limite K-Pg. A fauna associada inclui Tyrannosaurus rex, Triceratops horridus e prorsus, Torosaurus, Edmontosaurus annectens, Anzu wyliei, Pachycephalosaurus, Ankylosaurus magniventris, Thescelosaurus, Ornithomimus, Acheroraptor temertyorum (dromeossaurídeo menor) e troodontídeos como Pectinodon, além de crocodilianos (Borealosuchus), tartarugas (incluindo a trionichídea Axestemys splendida, crucial para a controvérsia das 'fúrculas' de Dakotaraptor), mamíferos, répteis escamados e aves.
Galeria de imagens
















Reconstituição em vida de Dakotaraptor steini com plumagem completa sobre o antebraço, inferida a partir das cerca de 15 papilas ulnares (quill knobs) preservadas na ulna do holótipo.
Wikimedia Commons
Ecologia e comportamento
Habitat
Planícies costeiras quentes e úmidas do interior norte-americano durante o Maastrichtiano tardio, drenadas por rios meandrantes que atravessavam florestas subtropicais dominadas por coníferas (Taxodium, araucariáceas), palmeiras (Arecaceae), cicadáceas, angiospermas de folha larga (Dryophyllum) e pteridófitas. A costa do epicontinental Western Interior Seaway ainda existia durante a deposição do Hell Creek, mas já recuava rapidamente. A unidade inclui canais fluviais ativos, planícies de inundação com solos hidromórficos, pântanos e lagoas efêmeras. A fauna inclui Tyrannosaurus rex como predador dominante, Triceratops e Torosaurus entre os ceratopsídeos, Edmontosaurus annectens entre os hadrossaurídeos, Ankylosaurus e Denversaurus como anquilossauros, Pachycephalosaurus e Thescelosaurus, e, entre os terópodes pequenos, Acheroraptor, Anzu, Ornithomimus e Troodon / Pectinodon. Dakotaraptor convive com toda essa fauna nas últimas centenas de milhares de anos do Cretáceo.
Alimentação
Predador ativo, carnívoro estrito. O tamanho intermediário entre o jovem Tyrannosaurus rex e pequenos terópodes como Acheroraptor sugere que Dakotaraptor ocupava um nicho de mesopredador, possivelmente explorando presas que estavam fora do alcance de tiranossaurídeos juvenis mas em vantagem frente a Acheroraptor e Troodon. Presas prováveis incluíam Thescelosaurus, Pachycephalosaurus juvenis, ornitomimossauros como Ornithomimus e hadrossaurídeos jovens. A garra de 24 cm do segundo dedo do pé, a famosa sickle claw, e os membros posteriores esguios, com tíbia muito longa, sugerem tanto estratégia de predação ativa em terreno aberto quanto capacidade de segurar e imobilizar presas, como hipotetizado por Fowler et al. (2011) para Deinonychus.
Comportamento e sentidos
Não há trilhos ou sítios inequivocamente atribuíveis a Dakotaraptor, e o hipodigma reduzido pós-corrigendum limita inferências diretas. Por analogia com outros eudromeossauros, é possível um comportamento de caçada solitária ou em pares esporádicos, com corrida rápida em terreno aberto e ataque com a sickle claw para imobilizar a presa. A presença de papilas ulnares indicando plumagem no antebraço sugere que exibição visual, abrigo sobre ninhos e termorregulação eram papéis importantes das penas no animal adulto, independentemente de qualquer capacidade de voo, inexistente dado o porte.
Fisiologia e crescimento
Dakotaraptor foi um dromeossaurídeo de porte excepcional e anatomia incomumente esguia entre os grandes representantes da família. A tíbia de 678 mm é a mais longa conhecida em Dromaeosauridae, indicando membros posteriores proporcionalmente longos e potencialmente maior capacidade cursorial que Utahraptor (robusto) e Achillobator (maciço). As papilas ulnares atestam penas grandes no antebraço, homólogas às secundárias de aves modernas. A histologia óssea do táxon não foi publicada em detalhe, o que é particularmente sensível dada a controvérsia de identificação do material, mas Burnham e colegas anunciaram, em apresentações de congresso pós-2015, que a microanatomia do fêmur e da ulna é compatível com dromeossaurídeos.
Paleogeografia
Configuração continental

Ron Blakey · CC BY 3.0 · Cretáceo, ~90 Ma
Durante o Maastrichtiano tardio (~67–66 Ma), Dakotaraptor steini habitava a Laramídia, a metade ocidental do que hoje é a América do Norte, separada pelo Mar Interior do Oeste (Western Interior Seaway), um mar raso que dividia o continente ao meio. Os continentes estavam em posições muito diferentes das atuais: a Índia viajava em direção à Ásia, a Antártida ainda estava conectada à Austrália, e a América do Sul era uma ilha separada.
Inventário de Ossos
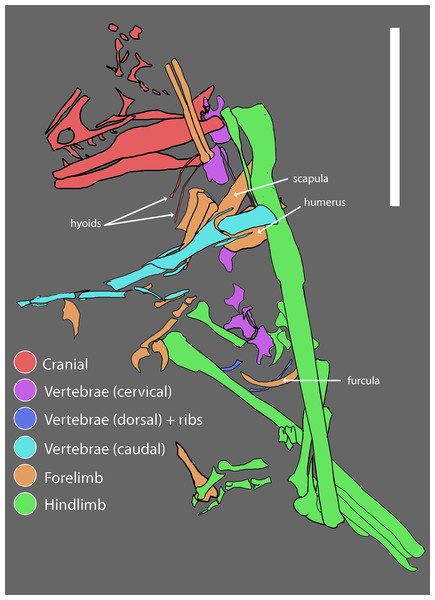
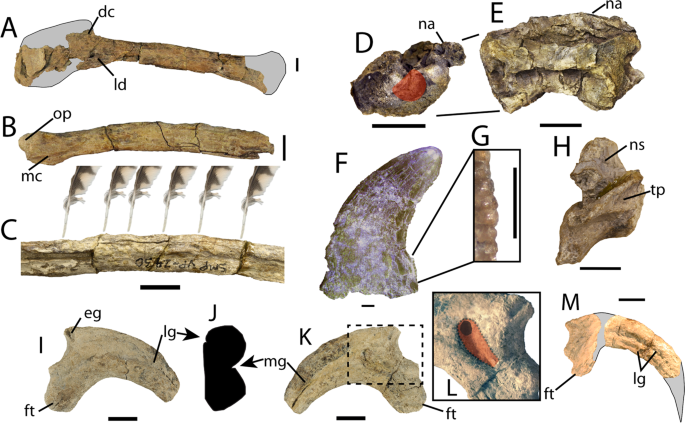
O holótipo PBMNH.P.10.113.T, após o corrigendum de DePalma et al. (2016), é composto basicamente por membros anteriores, membros posteriores e vértebras caudais de um único indivíduo adulto, sem crânio nem maior parte da coluna axial. A tíbia esquerda, preservada completa com 678 mm, é a mais longa conhecida em qualquer dromeossaurídeo. A ulna preserva cerca de 15 papilas ulnares, as famosas quill knobs que indicam a inserção direta de penas grandes na borda caudal do antebraço, semelhantes às descritas para Velociraptor por Turner e colegas em 2007. Um dos aspectos mais discutidos da taxonomia do animal é a inacessibilidade do material-tipo: o Palm Beach Museum of Natural History é descrito por vários autores como coleção privada, o que limita a reanálise independente, e Cau (2023 a 2024) argumenta, ainda em blog, que o material pode ser uma quimera.
Estruturas encontradas
Estruturas inferidas
Literatura Científica
15 artigos em ordem cronológica — do artigo de descrição original até pesquisas recentes.
Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana
Ostrom, J.H. · Bulletin of the Peabody Museum of Natural History 30: 1 to 165
Monografia fundadora dos Dromaeosauridae modernos. A descrição de Deinonychus antirrhopus, com sua 'garra terrível' do segundo dedo do pé e a semelhança manual com aves, reavivou a hipótese de ancestralidade dinossauriana das aves e é base indispensável para interpretar a anatomia de Dakotaraptor.


A large dromaeosaur (Theropoda) from the Lower Cretaceous of eastern Utah
Kirkland, J.I., Burge, D. e Gaston, R. · Hunteria 2(10): 1 to 16
Descrição original de Utahraptor ostrommaysorum, o gigante dromeossaurídeo do Cretáceo Inferior da Formação Cedar Mountain, Utah. Com cerca de 7 metros e ossos do crânio 2,5 vezes maiores que os de Deinonychus, foi por duas décadas o maior dromeossaurídeo conhecido, até ser equiparado por Dakotaraptor em 2015.


A new maniraptoran theropod, Achillobator giganticus (Dromaeosauridae), from the Upper Cretaceous of Burkhant, Mongolia
Perle, A., Norell, M.A. e Clark, J.M. · Contributions of the Mongolian-American Paleontological Project
Descrição de Achillobator giganticus, dromeossaurídeo mongol do Cretáceo Superior (~96 a 89 Ma), com esqueleto robusto, pelve primitiva com púbis verticalizado e tamanho entre 4,5 e 5 metros. Referência-chave para entender a diversidade de grandes eudromeossauros e para comparar a ecologia de um predador maciço com o porte esguio de Dakotaraptor.

Feather quill knobs in the dinosaur Velociraptor
Turner, A.H., Makovicky, P.J. e Norell, M.A. · Science 317(5845): 1721
Primeira demonstração direta de quill knobs (papilas ulnares) em um dromeossaurídeo não aviano, com base na ulna de Velociraptor mongoliensis (IGM 100/981). A feição é idêntica à observada por DePalma et al. (2015) em Dakotaraptor, e é a base da interpretação de que raptores gigantes do Maastrichtiano tinham antebraços emplumados apesar do porte incompatível com voo.
A microraptorine (Dinosauria-Dromaeosauridae) from the Late Cretaceous of North America
Longrich, N.R. e Currie, P.J. · Proceedings of the National Academy of Sciences 106(13): 5002 to 5007
Descrição de Hesperonychus elizabethae, o menor dromeossaurídeo não aviano conhecido da América do Norte, da Formação Dinosaur Park (Campaniano, Alberta). Amplia o registro temporal de Microraptorinae em 45 milhões de anos e estabelece o pano de fundo para discutir a diversidade de tamanho dos dromeossaurídeos norte-americanos do Cretáceo Superior, da qual Dakotaraptor representa o extremo oposto.
A review of dromaeosaurid systematics and paravian phylogeny
Turner, A.H., Makovicky, P.J. e Norell, M.A. · Bulletin of the American Museum of Natural History 371: 1 to 206
Revisão monográfica da sistemática de Dromaeosauridae e da filogenia de Paraves. Define Eudromaeosauria como o clado que reúne Dromaeosaurinae, Velociraptorinae e Saurornitholestinae, o quadro taxonômico dentro do qual Dakotaraptor foi originalmente posicionado em 2015. Base matricial para análises subsequentes de Jasinski (2020) e Currie e Evans (2019).
A new dromaeosaurid (Dinosauria: Theropoda) with Asian affinities from the latest Cretaceous of North America
Evans, D.C., Larson, D.W. e Currie, P.J. · Naturwissenschaften 100(11): 1041 to 1049
Descrição de Acheroraptor temertyorum a partir de material craniano da Formação Hell Creek de Montana. Até então considerado o único dromeossaurídeo de Hell Creek, foi acompanhado dois anos depois por Dakotaraptor, demonstrando que a formação abrigava pelo menos dois dromeossaurídeos contemporâneos em classes de tamanho muito diferentes.


The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation
DePalma, R.A., Burnham, D.A., Martin, L.D., Larson, P.L. e Bakker, R.T. · Paleontological Contributions 14: 1 to 16
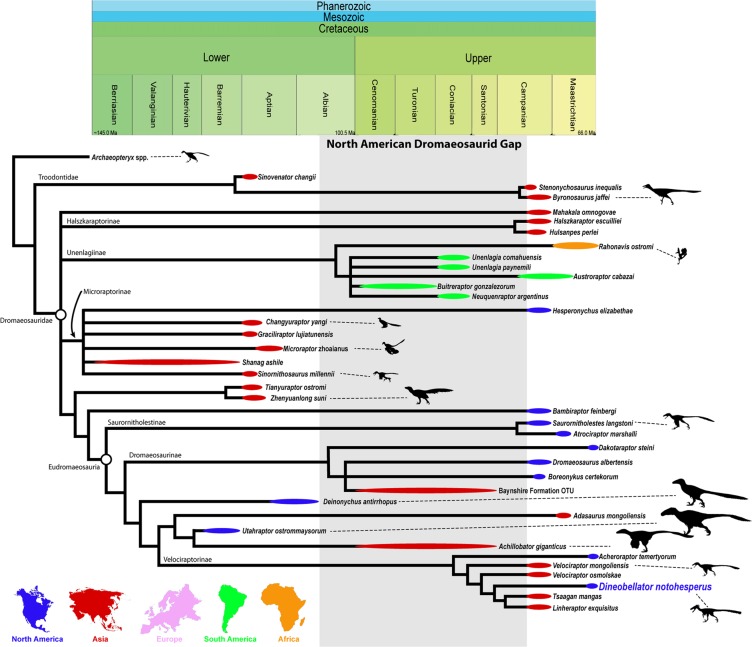
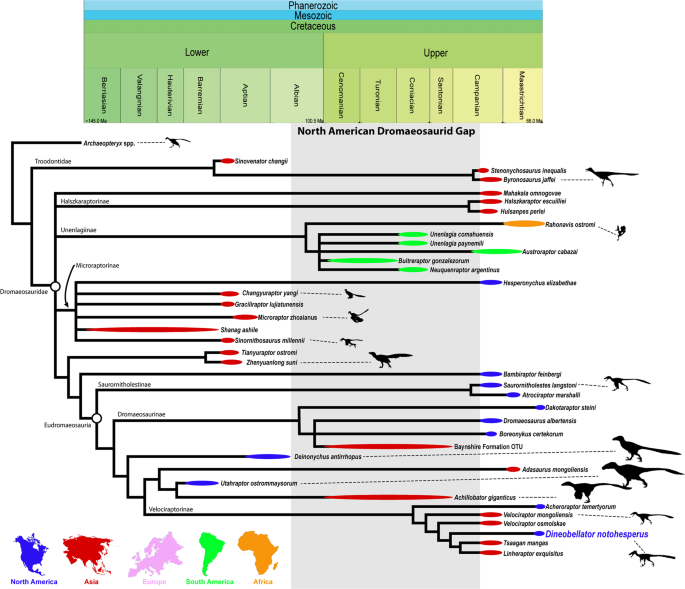
Descrição original de Dakotaraptor steini com base no holótipo PBMNH.P.10.113.T, coletado por Robert DePalma em 2005 na parte superior da Formação Hell Creek, Harding County, South Dakota, a menos de 20 metros abaixo do limite Cretáceo-Paleógeno. Os autores destacam: tamanho de cerca de 5,5 metros e massa estimada em cerca de 350 kg, tíbia de 678 mm (a mais longa conhecida em Dromaeosauridae), cerca de 15 papilas ulnares indicando penas grandes no antebraço e três elementos descritos como fúrculas. A análise filogenética posicionou Dakotaraptor como eudromeossauro próximo a Dromaeosaurus. Esta descrição é a referência primária e deve ser citada em conjunto com o corrigendum de 2016 e a crítica de Arbour et al. (2016).
The furculae of the dromaeosaurid dinosaur Dakotaraptor steini are trionychid turtle entoplastra
Arbour, V.M., Zanno, L.E., Larson, D.W., Evans, D.C. e Sues, H.-D. · PeerJ 4: e1691
Correção taxonômica crucial. Os autores demonstram que as três 'fúrculas' atribuídas a Dakotaraptor steini no artigo original, incluindo a peça incorporada ao holótipo, não são ossos de terópode, mas sim entoplastros (placas centrais do plastrão) da tartaruga trionichídea Axestemys splendida, comum na Formação Hell Creek. A identificação se baseia na forma achatada em V/U, na ausência de cabeça epicleidial distinta, na microanatomia esponjosa sem padrão laminado e na congruência perfeita com Axestemys conhecida do mesmo sítio. O artigo obrigou os autores originais a emitir o corrigendum de 2016.


Corrigendum to: The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation
DePalma, R.A., Burnham, D.A., Martin, L.D., Larson, P.L. e Bakker, R.T. · Paleontological Contributions 16: 1 to 2
Corrigendum formal dos autores originais, publicado em 2 de dezembro de 2016. Aceita a reidentificação de Arbour et al. (2016) das 'fúrculas' como entoplastros de Axestemys splendida. Exclui do hipodigma de Dakotaraptor a fúrcula anteriormente atribuída ao holótipo e os espécimes referidos KUVP 152429 (exclusivamente material de tartaruga) e NCSM 13170. O hipodigma reduzido permanece como base para toda a discussão posterior sobre o táxon.
Cranial anatomy of new specimens of Saurornitholestes langstoni (Dinosauria, Theropoda, Dromaeosauridae) from the Dinosaur Park Formation (Campanian) of Alberta
Currie, P.J. e Evans, D.C. · The Anatomical Record 303(4): 691 to 715
Descrição da anatomia craniana de um esqueleto quase completo de Saurornitholestes langstoni da Formação Dinosaur Park (Campaniano, Alberta). Inclui nova análise filogenética em que Dakotaraptor é mantido dentro de Eudromaeosauria, mas em posição sensível à remoção de caracteres não verificáveis, em linha com as críticas posteriores de Cau.


A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight
Hartman, S., Mortimer, M., Wahl, W.R., Lomax, D.R., Lippincott, J. e Lovelace, D.M. · PeerJ 7: e7247
Descrição do troodontídeo Hesperornithoides miessleri e grande análise filogenética de Paraves. No esquema de Hartman e colegas, Dakotaraptor foi recuperado dentro de Unenlagiidae (dromeossaurídeos sul-americanos e antárticos) em vez de Eudromaeosauria. Essa instabilidade topológica é uma das principais razões pelas quais a posição de Dakotaraptor permanece contestada.

New Dromaeosaurid Dinosaur (Theropoda, Dromaeosauridae) from New Mexico and biodiversity of dromaeosaurids at the end of the Cretaceous
Jasinski, S.E., Sullivan, R.M. e Dodson, P. · Scientific Reports 10: 5105
Descrição de Dineobellator notohesperus, primeiro dromeossaurídeo diagnóstico do final do Cretáceo no sul dos Estados Unidos (New Mexico). A análise filogenética recupera Eudromaeosauria com Saurornitholestinae, Dromaeosaurinae e Velociraptorinae, com Dakotaraptor e Acheroraptor entre os terminais maastrichtianos. Este é o cladograma publicado mais citado para posicionar Dakotaraptor em árvores recentes.
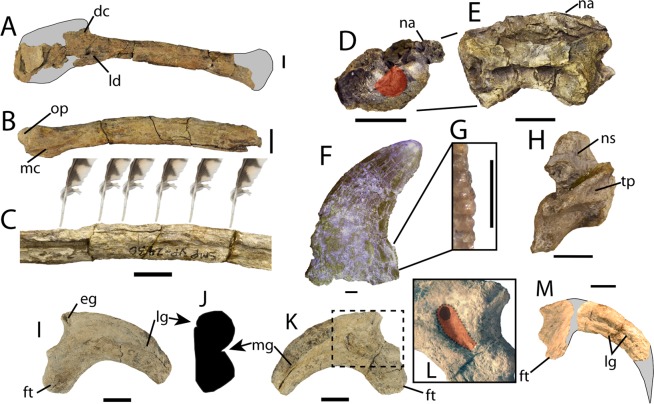
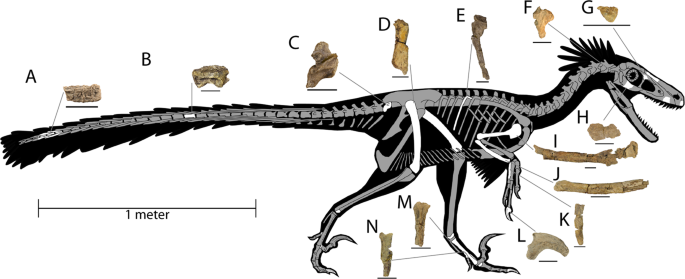
Osteology and reassessment of Dineobellator notohesperus, a southern eudromaeosaur (Theropoda: Dromaeosauridae: Eudromaeosauria) from the latest Cretaceous of New Mexico
Jasinski, S.E. · The Anatomical Record 306(7): 1641 to 1708
Monografia osteológica de Dineobellator notohesperus com revisão ampla de Eudromaeosauria. Discute a diversidade corporal dos dromeossaurídeos maastrichtianos, posicionando Dineobellator como eudromeossauro de porte médio e Dakotaraptor como extremo de tamanho do mesmo clado. Reforça, com mais caracteres, que Dakotaraptor é parte de Eudromaeosauria, embora reconheça a instabilidade topológica apontada por Hartman et al. (2019) e por Cau.
Calibration of chron C29r: new high-precision geochronologic and paleomagnetic constraints from the Hell Creek region, Montana
Sprain, C.J., Renne, P.R., Clemens, W.A. e Wilson, G.P. · GSA Bulletin 130(9-10): 1615 to 1644
Calibração geocronológica de alta precisão (U-Pb CA-TIMS em zircão e 40Ar/39Ar) da Formação Hell Creek em Montana. Estabelece o limite K-Pg em 66,043 ± 0,010 Ma e restringe a duração da deposição do topo do Hell Creek a menos de 1 milhão de anos. Contexto essencial para afirmar que Dakotaraptor viveu nos últimos 100 a 400 mil anos do Cretáceo, a menos de 20 m do limite K-Pg no sítio holótipo.


Espécimes famosos em museus
PBMNH.P.10.113.T (holótipo de Dakotaraptor steini)
Palm Beach Museum of Natural History, Flórida, EUA
Holótipo de Dakotaraptor steini, coletado em 2005 em um canal fluvial da parte superior da Formação Hell Creek, Harding County, South Dakota, a menos de 20 metros abaixo do limite Cretáceo-Paleógeno. O material foi depositado no Palm Beach Museum of Natural History (Flórida), descrito por vários autores como coleção privada, o que tem gerado críticas quanto à acessibilidade para reanálise independente. Após o corrigendum de DePalma et al. (2016), a fúrcula incorporada ao holótipo foi removida do hipodigma por ter sido reidentificada como entoplastro da tartaruga Axestemys splendida por Arbour et al. (2016).
PBMNH.P.10.115.T e PBMNH.P.10.118.T (referidos)
Palm Beach Museum of Natural History, Flórida, EUA
Espécimes referidos no artigo original de 2015 que permaneceram no hipodigma após o corrigendum de 2016. São fragmentos e ossos isolados cuja associação a Dakotaraptor é baseada em critérios de tamanho e morfologia compatíveis com o holótipo. Ainda assim, a análise de Cau (2023 a 2024) levanta dúvidas sobre a homogeneidade do conjunto.
KUVP 156045 (referido)
University of Kansas Museum of Natural History (KU Biodiversity Institute), Lawrence, Kansas, EUA
Um dos espécimes referidos originalmente ao táxon e mantidos após o corrigendum. Importante por estar em coleção pública acessível (KU), o que possibilita reavaliações independentes. O espécime KUVP 152429, também referido em 2015, foi excluído pelo corrigendum de 2016 por consistir exclusivamente em material de tartaruga.
Réplicas e montagens reconstruídas
Palm Beach Museum of Natural History (Flórida) e exposições itinerantes
Réplicas esqueléticas reconstruídas com material do holótipo e casts de Utahraptor e Achillobator foram exibidas em eventos educativos. Nenhuma montagem esquelética permanente e oficial de Dakotaraptor foi instalada em museu público de grande porte até 2026.
Classificação
Descoberta
Curiosidade
A parte mais famosa de Dakotaraptor talvez não seja a garra em foice de 24 cm, nem as papilas ulnares que mostram plumagem em um animal grande demais para voar: é a 'fúrcula que não era fúrcula'. Em 2015, DePalma e colegas descreveram três ossos em forma de U como fúrculas de Dakotaraptor, uma delas parte do holótipo. Em fevereiro de 2016, Arbour, Zanno, Larson, Evans e Sues mostraram que esses ossos são, na verdade, entoplastros (placas centrais do plastrão) da tartaruga trionichídea Axestemys splendida, comum na Formação Hell Creek. Os autores originais publicaram um corrigendum formal em dezembro de 2016 aceitando a correção. O episódio virou caso de estudo em paleontologia sobre a importância de pareceres independentes e acesso ao material-tipo, e é parte inseparável da identidade científica do táxon.