Tyrannosauroidea
A Família dos Reis
Os tiranossauros dominaram a Laurásia (América do Norte e Ásia) nos últimos 30 milhões de anos do Cretáceo. Começaram pequenos e emplumados: Yutyrannus, descoberto em 2012 na China, tinha 9 metros e pelugem filamentosa cobrindo o corpo. Só nos últimos 15 milhões de anos do Mesozoico evoluíram para o gigantismo dos tiranossaurídeos clássicos, com crânio maciço, dentes em forma de banana e braços de dois dígitos reduzidos. Esta árvore reúne as oito espécies de tiranossauro na nossa base, organizadas em dois grandes ramos: Albertosaurinae (predadores mais esbeltos e rápidos) e Tyrannosaurinae (carcadores robustos, incluindo T. rex e Tarbossauro).
Comparativo
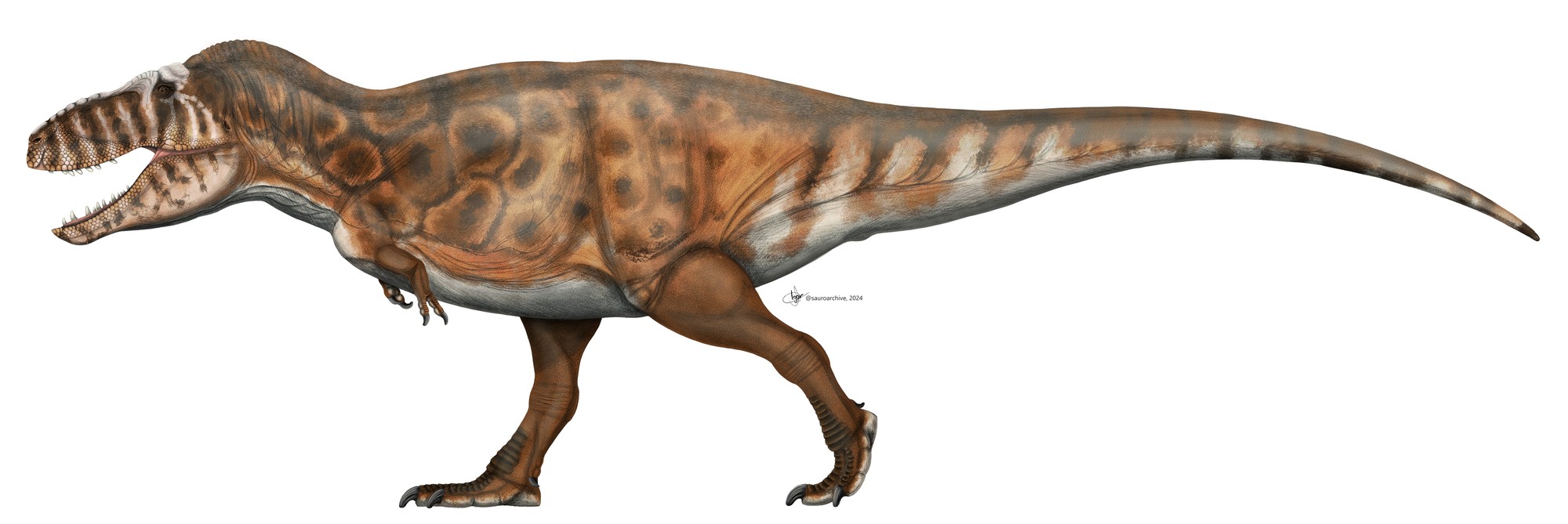
T. rex e Tarbossauro: o que é igual e o que é diferente?
Tarbosaurus bataar, da Mongólia, e Tyrannosaurus rex, da América do Norte, são os dois maiores tiranossauros já encontrados. Viveram quase no mesmo tempo geológico, em continentes separados por um mar raso. Por décadas, Tarbossauro foi classificado como uma espécie de T. rex ("Tyrannosaurus bataar"). Hoje a maioria dos paleontólogos os separa em gêneros distintos, mas as semelhanças continuam marcantes: ambos têm crânio maciço, dentes em banana, braços vestigiais de dois dígitos e ocupavam o mesmo nicho ecológico. As diferenças aparecem quando se olha com lupa.

| Característica | Tyrannosaurus rex | Tarbosaurus bataar |
|---|---|---|
| Idade | 68–66 Ma (Maastrichtiano) | 70–68 Ma (Maastrichtiano inicial) |
| Onde viveu | Laurásia ocidental (EUA, Canadá) | Laurásia oriental (Mongólia, China) |
| Comprimento | até 12 m | até 10 m |
| Peso | até 8 toneladas | até 5 toneladas |
| Crânio | 1,5 m, largo atrás, reforçado para mordida de esmagamento | 1,3 m, mais estreito e alongado, com ossos fusionados |
| Visão | Binocular forte: campo de sobreposição de 55°, entre os maiores de qualquer dinossauro | Binocular reduzida: órbitas mais laterais, menos sobreposição |
| Braços | Dois dígitos, ~1 m de comprimento | Dois dígitos, proporcionalmente menores que os do T. rex |
| Força de mordida | A maior já medida em um animal terrestre: ~35.000 N | Menor em termos absolutos, mas ainda capaz de esmagar ossos |
| Presas principais | Triceratops, Edmontosaurus, Ankylosaurus | Saurolophus, Therizinosaurus, saurópodes |
| Ambiente | Florestas costeiras úmidas, planícies aluviais | Planícies semi-áridas, ambientes fluviais sazonais |
| Parentesco | Ambos no clado Tyrannosaurini (dentro de Tyrannosaurinae). São os parentes vivos mais próximos já encontrados. | |
Resumo: irmãos geográficos. Tarbossauro é a versão asiática, um pouco menor e mais leve, adaptada a ambientes mais secos. T. rex é a versão norte-americana, mais pesado, com visão estereoscópica superior e a mordida mais potente já registrada. Os dois ocupam o topo da cadeia alimentar de seus continentes no último suspiro do Cretáceo.
Como dois primos foram parar em lados opostos do planeta?
No fim do Cretáceo, T. rex e Tarbossauro ocupavam nichos quase idênticos, ambos predadores apex de 8 a 12 metros, em continentes separados por milhares de quilômetros de oceano. Como dois parentes tão próximos acabaram em pontas opostas do mundo?
A resposta mais recente vem de um estudo de Morrison e colaboradores publicado na revista Royal Society Open Science em maio de 2025. Reconstruindo a biogeografia dos tiranossauros via modelagem estatística da filogenia, os autores concluíram que o ancestral direto do clado Tyrannosaurus–Tarbosaurus estava presente em ambos os continentes ao mesmo tempo, na Ásia e em Laramídia. Ou seja, antes de T. rex e Tarbossauro existirem como espécies, a população ancestral já havia se espalhado pelos dois lados do planeta, atravessando do Velho Mundo para o Novo.
O evento de dispersão ocorreu no fim do Campaniano e início do Maastrichtiano, entre cerca de 73 e 69 Ma. Foi a partir dessa população ancestral bicontinental que os dois gêneros se diferenciaram: o gênero Tyrannosaurus, segundo a análise, é endêmico de Laramídia, embora descenda de imigrantes asiáticos.

A ponte entre os dois continentes era Beríngia, uma vasta planície baixa e emersa que conectava a Sibéria oriental ao Alasca, larga de até 1.500 km, coberta por florestas temperadas quentes. No Cretáceo Final não existiam calotas polares, e o clima do extremo norte era suave o bastante para sustentar florestas e manadas inteiras de dinossauros. Sempre que o nível global do mar baixava, algo que aconteceu em pulsos sucessivos durante o Campaniano e o Maastrichtiano, Beríngia emergia em larga escala e abria um corredor biogeográfico que grandes vertebrados podiam atravessar caminhando. Morrison e colegas documentaram múltiplos eventos de dispersão de pantiranossaurianos pela ponte terrestre em direções opostas ao longo do Cretáceo Final.
Os mares epicontinentais, como o Mar Interior Ocidental que dividia a América do Norte em Laramídia, a oeste, e Apalachia, a leste, ou o Estreito de Turgai que separava a Europa da Ásia, eram barreiras reais ao movimento dentro de cada continente. Mas os tiranossauros nunca precisaram atravessá-los diretamente. Seguiam as margens e faziam a travessia intercontinental somente pelo norte, pela ponte terrestre de Beríngia. Na prática, a viagem de um Tarbossauro mongol até o território de um T. rex em Montana era uma caminhada de dezenas de milhares de quilômetros pela borda do Ártico.
Por que ficaram gigantes?
A mesma pesquisa aponta uma causa ecológica para o gigantismo dos tiranossaurinos. Os carcarodontossaurídeos, grupo de predadores apex que incluía o Giganotossauro e o Carcharodontosaurus, declinaram e desapareceram durante o Campaniano. Sua extinção abriu uma vaga ecológica, o nicho de predador de topo com presa de grande porte, que os tiranossaurinos preencheram rapidamente. Em alguns milhões de anos, passaram de carnívoros medianos para colossos de 8 toneladas. A associação estatística com mudanças climáticas detectada no estudo sugere que resfriamentos regionais também podem ter favorecido o aumento de massa corporal nessas linhagens.
O resultado: duas linhagens irmãs evoluindo em paralelo em ambientes diferentes. Tarbossauro adaptou-se às planícies semi-áridas da Ásia Central, caçando saurópodes titanossauros e terizinossauros gigantes. T. rex herdou as florestas úmidas e planícies aluviais de Laramídia, caçando ceratópsios e hadrossauros. Ambos coexistiram por cerca de 4 milhões de anos, até que o impacto do meteoro em Chicxulub, há 66 milhões de anos, tirou os dois do palco ao mesmo tempo.
Fonte: Morrison, C., Pittman, M., Nurgaliyev, B., & Salguero-Gómez, R. (2025). Rise of the king: Gondwanan origins and evolution of megaraptoran dinosaurs. Royal Society Open Science, 12(5), 242238. doi.org/10.1098/rsos.242238
Dromaeosauridae
Os Raptores Emplumados
Os dromeossaurídeos são os dinossauros não-aviários mais próximos das aves modernas. Todos tinham penas (confirmadas por fósseis em múltiplos membros do clado), garra falciforme elevada no segundo dedo, cauda rígida para equilíbrio em manobras ágeis e cérebro relativamente grande. Na nossa base estão cinco espécies que mostram a diversidade do grupo: do Microraptor minúsculo com quatro asas até o Utahraptor de seis metros. Esqueça o Velociraptor do cinema: o real tinha o tamanho de um peru.
Ornithischia
Os Herbívoros Ornamentados
Ornithischia é um dos dois grandes ramos dos dinossauros. Todos eram herbívoros e desenvolveram os ornamentos mais extravagantes do Mesozoico: placas dorsais (estegossauros), blindagem corporal completa com clube caudal (anquilossauros), cristas cranianas ocas para vocalização (hadrossauros), cúpulas cranianas de 25 cm (paquicefalossauros) e chifres com frila (ceratópsios). Esta árvore reúne todos os ornitísquios da nossa base, organizados em três grandes sub-clados: Thyreophora (os blindados), Ornithopoda (os herbívoros de mandíbula lateral) e Marginocephalia (os ornamentados de crânio).
Stegosauria
As Placas Dorsais
Os estegossauros surgiram no Jurássico Médio e viveram até o Cretáceo Inicial, com duas fileiras alternadas de placas ou espinhos ao longo do dorso e espinhos caudais, chamados de thagomizer, usados em defesa ativa. Diferenciam-se por crânio pequeno e alongado, dentes simples para vegetação mole e quadris mais altos que os ombros. Huayangosaurus, o mais basal do grupo, ainda tinha dentes na frente do focinho; os Stegosauridae derivados perderam esses dentes e desenvolveram placas cada vez mais elaboradas. Estudos mostram que as placas de Stegosaurus eram ricamente vascularizadas, sugerindo função termorreguladora ou de exibição, não armadura.
Ankylosauria
Os Tanques Vivos
Os anquilossauros eram os dinossauros mais blindados: osteodermos (placas ósseas embutidas na pele) cobriam todo o corpo, inclusive as pálpebras em algumas espécies. O grupo se divide em dois grandes ramos. Nodosauridae (inclui Borealopelta, Edmontonia, Sauropelta) tinham armadura alinhada com espinhos laterais proeminentes, sem clube caudal. Ankylosauridae (inclui Ankylosaurus, Euoplocephalus, Pinacosaurus, Zuul) desenvolveram um clube caudal maciço de osso fundido, arma ativa capaz de quebrar as tíbias de um T. rex, segundo estudos biomecânicos recentes. Minmi, da Austrália, pertence a uma linhagem basal chamada Parankylosauria, separada dos dois ramos principais há mais de 150 milhões de anos.
Ceratopsia
Os Chifres e as Frilas
Os ceratópsios começaram pequenos e bípedes, como Psittacosaurus, e evoluíram para os grandes herbívoros quadrúpedes com chifres e frilas elaboradas do Cretáceo Final da Laurásia. O grupo se divide em duas linhagens basais (Psittacosauridae e Protoceratopsidae, sem chifres verdadeiros) e na grande família Ceratopsidae, que desenvolveu chifres ósseos reais. Dentro de Ceratopsidae há dois ramos: Centrosaurinae (frila curta, chifre nasal proeminente, membros como Nasutoceratops, Pachyrhinosaurus, Styracosaurus) e Chasmosaurinae (frila longa, chifres supraorbitais longos, membros como Chasmosaurus, Kosmoceratops, Torosaurus e o icônico Triceratops). O crânio de Triceratops ultrapassa 2,5 m, entre os maiores de qualquer animal terrestre.