Paleobotânica
A Evolução das Plantas
De florestas sem flores dominadas por coníferas gigantes a campos de gramíneas que cobrem 40% da Terra. A história das plantas ao longo do Mesozoico e Cenozoico é tão dramática quanto a dos dinossauros, e as duas estão profundamente entrelaçadas.
Florestas sem flores
O mundo vegetal dos dinossauros era radicalmente diferente do nosso. Sem gramíneas, sem flores, sem frutos.
Durante o Triássico e Jurássico, o dossel florestal era dominado por coníferas gigantes das famílias Araucariaceae e Cheirolepidiaceae, que alcançavam 30 a 60 metros. Abaixo, cicadáceas, bennettitales e ginkgoales formavam o estrato intermediário, e o solo era coberto por samambaias, musgos e cavalinhas. Não existiam gramíneas, flores ou frutos. Esse cenário persistiu por mais de 100 milhões de anos.
A dieta dos saurópodes dependia dessas plantas. Análises de isótopos de carbono em esmalte dentário (Formação Morrison, EUA) indicam que Brachiosaurus alcançava o dossel das araucárias, enquanto Diplodocus pastava samambaias no solo. Testes laboratoriais (Hummel et al., 2008) mostraram que folhagens de Araucaria e cavalinhas produzem energia comparável a forragens modernas, viabilizando o gigantismo desses animais.
Sítios fósseis notáveis
Floresta Petrificada do Arizona (~225 Ma): troncos silicificados de coníferas do Triássico Superior.
Cerro Cuadrado, Patagônia (~160 Ma): cones de Araucaria mirabilis com preservação celular e embriões intactos.
Skåne, Suécia: samambaias jurássicas com núcleos celulares e cromossomos preservados (Bomfleur et al., 2014).


Floresta jurássica vs. moderna
As "flores" antes das flores: Bennettitales
Uma ordem extinta que desenvolveu estruturas semelhantes a flores de forma completamente independente das angiospermas. Um caso notável de evolução convergente.
As Bennettitales foram uma ordem de plantas gimnospérmicas que viveram por quase 170 milhões de anos. Externamente parecidas com as cicadáceas atuais, com troncos robustos e folhas pinadas, escondiam uma diferença crucial: seus órgãos reprodutivos. Algumas espécies, como Williamsonia e Cycadeoidea, desenvolveram estruturas semelhantes a flores, com brácteas petaloides e órgãos masculinos e femininos reunidos na mesma unidade reprodutiva.
O declínio das Bennettitales está intimamente ligado à ascensão das angiospermas. À medida que as plantas com flores se diversificaram a partir de ~100 Ma, conquistaram os nichos ecológicos antes ocupados pelas Bennettitales. Com taxas de crescimento mais rápidas e relações mutualísticas com polinizadores, as angiospermas levaram vantagem competitiva decisiva. As Bennettitales desapareceram no limite K-Pg (66 Ma), possivelmente já em declínio severo antes do impacto de Chicxulub.
Evolução convergente
A antiga "Hipótese Antófita" propunha que Bennettitales seriam ancestrais diretas das plantas com flores. Análises filogenéticas modernas refutaram essa ideia: Bennettitales são mais próximas de cícadas e ginkgos, não de angiospermas. A semelhança floral é puramente convergente, o que torna o grupo ainda mais fascinante.
Duas famílias
Williamsoniaceae
Troncos ramificados, "flores" expostas com brácteas petaloides. Visualmente semelhantes a flores de angiospermas.
Cycadeoidaceae
Troncos compactos em formato de barril. Estruturas reprodutivas embutidas na superfície, muitas vezes fechadas (cleistogamia).
O "mistério abominável": a revolução das angiospermas
Charles Darwin, em carta a Joseph Hooker de 1879, chamou o surgimento explosivo das plantas com flores de "mistério abominável". Em 30 milhões de anos, passaram de marginais a dominantes.
"The rapid development as far as we can judge of all the higher plants within recent geological times is an abominable mystery."
Charles Darwin, carta a Joseph Hooker, 22 de julho de 1879
Os registros fósseis mais antigos de angiospermas incluem grãos de pólen de ~136 milhões de anos encontrados em Israel, a planta aquática Montsechia vidalii (~130 Ma, Espanha) e Archaefructus sinensis (~125 Ma, China). Em apenas 30 milhões de anos, passaram de elementos marginais a dominantes nos ecossistemas terrestres, uma das transições ecológicas mais rápidas da história.
O impacto na fauna foi profundo. A diversificação dos dinossauros ornitísquios no Cretáceo Superior coincide com a dominância das angiospermas. Os hadrossaurídeos (dinossauros "bico de pato") desenvolveram baterias dentárias complexas, com centenas de dentes compactados formando superfícies de trituração para processar folhas duras e ricas em sílica das novas plantas.

Fósseis mais antigos
Grãos de pólen de angiospermas em Israel e Suíça
Montsechia vidalii, planta aquática da Espanha (Gomez et al., 2015)
Archaefructus sinensis, Formação Yixian, China (Sun et al., 2002)
Adaptações decisivas
Vasos no xilema: transporte de água mais eficiente que traqueídes de coníferas
Folhas largas: maior densidade de nervuras, maximizando a fotossíntese
Polinização por insetos: reprodução precisa e especiação acelerada
Frutos: dispersão de sementes por animais, colonizando novos habitats
Dupla fertilização: produz endosperma somente quando há fecundação, conservando energia
Flores e polinizadores: coevolução
A relação entre flores e insetos polinizadores é uma das parcerias mais bem-sucedidas da evolução. E começou no mundo dos dinossauros.
As primeiras evidências de polinização por insetos remontam a ~99 milhões de anos, preservadas em âmbar birmanês (Myanmar). Fósseis de tripes (Gymnopollisthrips minor) carregando grãos de pólen e besouros (Cretoparacucujus cycadophilus) associados a pólen de cicadáceas demonstram que a polinização entomófila surgiu antes mesmo da grande diversificação das angiospermas. Moscas de probóscide longa também aparecem nesse período.
A polinização por insetos acelerou dramaticamente a diversificação das angiospermas. Ao permitir isolamento reprodutivo e especiação rápida, cada par flor-polinizador podia divergir independentemente. Estudos com relógio molecular (Magallón et al., 2015) mostram que as principais linhagens de angiospermas (rosídeas, asterídeas) diversificaram-se entre 115 e 90 Ma, coincidindo com a diversificação dos polinizadores.
Progressão das síndromes de polinização
Besouros
Mais antiga; visitando cones de gimnospermas
Moscas
Probóscides longas no Cretáceo médio (~99 Ma)
Abelhas
Melittosphex burmensis (~100 Ma), a abelha mais antiga
Borboletas e mariposas
Mariposas com probóscide desde ~130 Ma; borboletas no Paleógeno
Evidência-chave: âmbar birmanês (~99 Ma)
Múltiplos grupos de insetos preservados carregando pólen: tripes, besouros, moscas. O âmbar de Myanmar é Cenomaniano e contém também uma das flores mais antigas preservadas em resina, Micropetasos burmensis, com estruturas que indicam polinização por insetos.
Dinossauros e angiospermas: coevolução ou coincidência?
As plantas com flor explodiram durante o Cretáceo, saindo de 0% para 85% da flora típica em cerca de 65 milhões de anos. Os dinossauros atingiram seu próprio pico de diversidade na mesma janela. Mike Benton, em uma revisão de 2024 sobre o "boom dos dinossauros no Cretáceo", examinou se os dois fenômenos estão ligados por causalidade ou se apenas coincidiram no tempo.
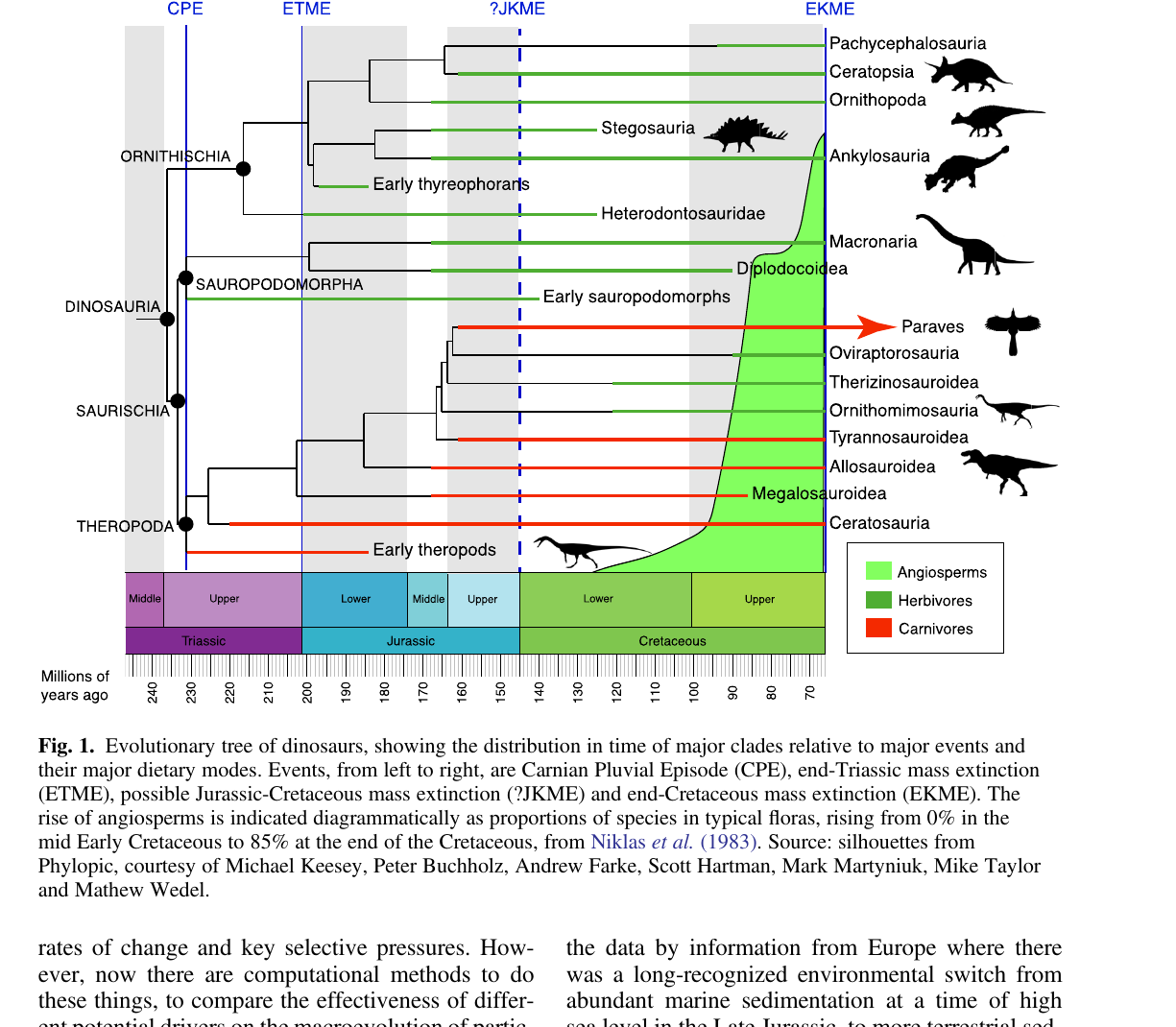
Angiospermas e gimnospermas são grupos-irmãos: a evidência filogenômica exige que a linhagem-caule das angiospermas tenha surgido ao mesmo tempo que as gimnospermas, no mais tardar no Triássico Superior (Li et al. 2019). Os fósseis mais antigos amplamente aceitos, porém, são pólens característicos do Valanginiano-Hauteriviano (140 a 130 Ma) e só aparecem no início do Cretáceo. Restos macroscópicos de angiospermas, incluindo flores, folhas, sementes e outros órgãos fossilizados, surgem a partir do Barremiano-Albiano (129 a 100 Ma).
Por que as angiospermas tiveram sucesso tão rápido? Benton lista as características únicas que as diferenciaram: dupla fertilização (o grão só vira semente depois que a fertilização está confirmada, economizando recursos), flor e coevolução com insetos polinizadores e, de forma decisiva, duas inovações do Cretáceo Superior na fisiologia foliar. A densidade estomática subiu de cerca de 100 para 1.000 poros por mm², e a densidade de nervuras foliares subiu de aproximadamente 4 para 9 mm de nervura por mm² de folha. As duas coisas são adaptações para absorção mais rápida de CO₂ e transporte mais rápido de água e nutrientes. Juntas, tornaram as angiospermas mais eficientes fotossinteticamente do que as gimnospermas que estavam substituindo.
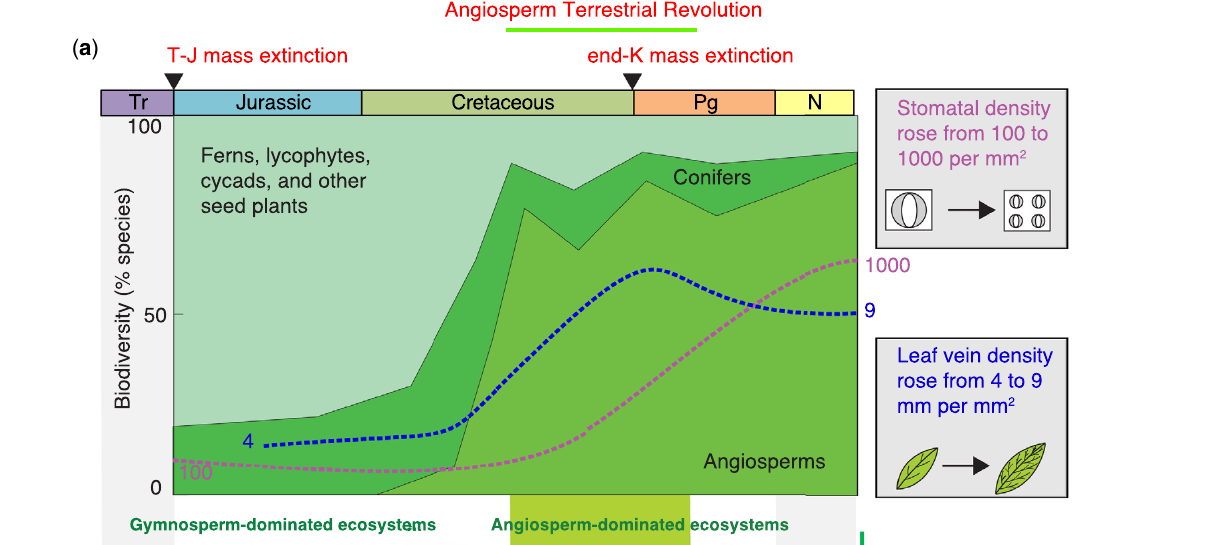
A Revolução Terrestre das Angiospermas (ATR)
Benton e colaboradores (2022) batizaram a expansão ecológica das plantas com flor de Revolução Terrestre das Angiospermas (Angiosperm Terrestrial Revolution, ATR) e a dataram entre 100 e 50 milhões de anos atrás, dividida em duas fases. A fase cretácea (100 a 66 Ma) é a que importa para a evolução dos dinossauros: as angiospermas passaram de presença marginal para domínio da flora típica em cerca de 34 milhões de anos. A segunda fase, no Paleoceno, depois do impacto do asteroide, marcou a origem das florestas tropicais. A travessia da marca de 50%, saindo do domínio gimnospérmico para o angiospérmico, acontece perto do limite Cenomaniano-Turoniano, por volta de 94 Ma.
Os dinossauros impulsionaram a revolução das angiospermas?
A hipótese clássica vem de Robert Bakker (1978, 1986): os saurópodes eram ramoneadores de altura, arrancavam folhas do dossel e não pastavam perto do chão. No início do Cretáceo, muitos clados de saurópodes entraram em declínio, e os novos herbívoros dominantes, os ornitópodes e os anquilossauros nodossaurídeos, passaram a comer rente ao solo. Teriam, portanto, pastado as mudas de crescimento lento das gimnospermas, abrindo espaço para as angiospermas, que cresciam mais rápido. No cenário de Bakker, o sucesso das angiospermas foi consequência de uma troca da guarda entre dinossauros.
Estudos posteriores questionaram a cronologia. Butler e colaboradores (2009, 2010) encontraram pouca evidência reproduzível de coevolução entre grupos de dinossauros e angiospermas ao testarem coincidências temporais e espaciais de ocorrência ao longo do registro cretáceo. E os saurópodes, na verdade, não sumiram no limite Jurássico-Cretáceo: continuaram representando 25 a 50% da diversidade herbívora total durante o Cretáceo Inferior (Barrett & Willis 2001). A "grande saída dos saurópodes" se revelou gradual e regional, sem abruptismo global.
A revisão de Benton vai além e compara as curvas de diversificação dos três grandes clados de dinossauros com a curva das angiospermas, usando riqueza genérica da Paleobiology Database. O padrão é sugestivo, mas não é estreito.
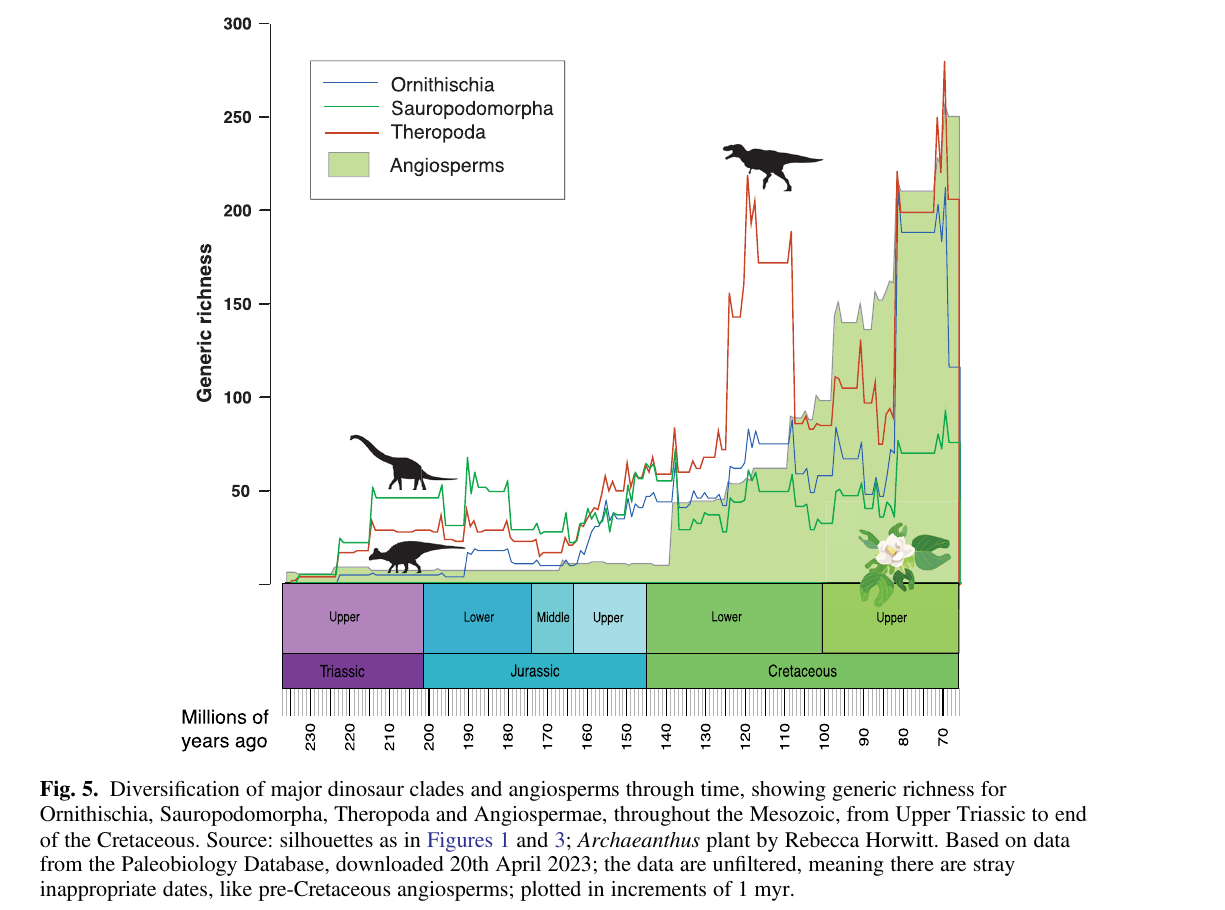
Dois pontos saltam aos olhos na Figura 5. A riqueza dos sauropodomorfos fica basicamente estável entre 40 e 80 gêneros durante todo o Cretáceo. Se as angiospermas estivessem remodelando o nicho herbívoro, a diversidade dos sauropodomorfos deveria se mover para cima ou para baixo em compasso com a virada florística. Não se move. Os ornitísquios, o clado que Bakker colocou no centro da sua história, só explodem em diversidade no finalzinho do Cretáceo, bem depois da ascensão das angiospermas. A curva dos terópodes correlaciona melhor com a das angiospermas do que qualquer das duas curvas de herbívoros, o que é difícil de explicar por pressão direta de herbivoria.
Evidência direta: o que os dinossauros efetivamente comiam?
Embora as curvas globais não mostrem coevolução estreita, conteúdos estomacais e coprólitos específicos não deixam dúvida: os dinossauros comiam angiospermas quando elas estavam disponíveis. Benton destaca três dos casos mais claros:
Anquilossauro Minmi
Cretáceo Inferior (Aptiano, cerca de 112 Ma) da Austrália. O conteúdo intestinal preservado era composto inteiramente por frutos de angiospermas (Molnar & Clifford 2000).
Coprólitos de hadrossauros
Campaniano de Montana. Biomarcadores moleculares específicos de angiospermas (Chin & Brassell 1994), junto com evidência de que os animais também comiam madeira de coníferas em decomposição (Chin 2007). Dieta mista.
Coprólitos de titanossauros
Maastrichtiano final da Índia. Continham fungos parasitas específicos de angiospermas, fitólitos de gramíneas e fitólitos de coníferas e palmeiras (Prasad et al. 2005). Os fitólitos de gramíneas empurraram a origem da família Poaceae para dentro da era dos dinossauros.
A conclusão de Benton
Os dinossauros comiam angiospermas, mas o descompasso temporal visível na Figura 5 pesa contra a ideia de que a herbivoria dos dinossauros tenha impulsionado a revolução das angiospermas, ou vice-versa. As angiospermas tiveram sucesso, principalmente, por méritos próprios: dupla fertilização, coevolução com polinizadores e, sobretudo, o salto do Cretáceo Superior em densidade estomática e de nervuras, que lhes deu vantagem de eficiência fotossintética sobre as gimnospermas que acabaram substituindo.
Fonte: Benton, M. J. (2024). The dinosaur boom in the Cretaceous. Geological Society, London, Special Publications, 544, 453-475. doi.org/10.1144/SP544-2023-70. Acesso aberto sob licença CC BY 4.0. Figuras 1, 4a (recortada da Figura 4) e 5 reproduzidas nos termos da licença.
O pico das samambaias: assinatura de catástrofe
Quando tudo morreu, as samambaias foram as primeiras a voltar. O "fern spike" é o sinal palinológico mais claro de colapso ecológico no registro fóssil.
Identificado pela primeira vez por Tschudy et al. (1984) em rochas da Formação Hell Creek (Montana, EUA), o "fern spike" consiste no aumento abrupto de esporos de samambaias no registro palinológico: de cerca de 25% para mais de 70 a 100% dos palinomorfos, logo acima da camada de irídio depositada pelo impacto de Chicxulub. Vajda e Bercovici (2014) confirmaram o padrão em escala global, documentando fern spikes equivalentes na Nova Zelândia e no Japão.
O fern spike no limite K-Pg durou entre 1.000 e 10.000 anos, um intervalo geológico brevíssimo que revela a escala da devastação. Fenômenos análogos foram registrados após a extinção do fim do Permiano (~252 Ma), com licófitas dominando o registro, e no fim do Triássico (~201 Ma). A recuperação completa de florestas diversificadas levou entre 100.000 e 1.000.000 de anos.

Por que samambaias dominam após catástrofes?
Esporos microscópicos: produzidos aos milhões, dispersos pelo vento, sem depender de polinizadores
Toleram luz reduzida: sobrevivem ao "inverno de impacto" causado pela poeira atmosférica
Colonização rápida: crescem em solos pobres, ácidos e recém-expostos
Sem dependência animal: reprodução totalmente independente de fauna
Sequência de recuperação pós-impacto
Colapso da vegetação, camada de irídio
Fern spike: samambaias dominam a paisagem
Retorno gradual de angiospermas e coníferas
Recuperação completa de florestas diversificadas
Análogos modernos: após a erupção do Krakatoa (1883) e do Monte Santa Helena (1980), samambaias estiveram entre as primeiras plantas a se estabelecer sobre as cinzas vulcânicas.
E nesse mundo de coníferas, flores e samambaias, uma família de plantas começava silenciosamente sua ascensão...
O surgimento da grama
De planta marginal no Cretáceo a bioma dominante no Mioceno: a trajetória das gramíneas ao longo de 100 milhões de anos.

Origem das gramíneas
Estimativas moleculares situam a origem da família Poaceae entre 80 e 100 milhões de anos atrás, ainda no período Cretáceo. O fóssil mais antigo de gramínea foi encontrado em âmbar de Myanmar (~100 Ma). As primeiras gramíneas eram plantas pequenas de sub-bosque, crescendo à sombra de coníferas, samambaias e angiospermas que dominavam a paisagem mesozoica.
Dinossauros comiam grama
Em 2005, Prasad e colegas publicaram na Science uma descoberta que abalou a paleontologia: fitólitos de pelo menos cinco tipos diferentes de gramíneas foram encontrados em coprólitos de titanossauros na Formação Lameta, Índia central. Saurópodes gigantes do Cretáceo Superior já incluíam gramíneas em sua dieta. As gramíneas não eram dominantes, mas faziam parte da vegetação disponível.
As gramíneas sobrevivem à extinção
O impacto de Chicxulub matou os dinossauros não aviários, mas as gramíneas atravessaram a catástrofe. Como plantas herbáceas com meristemas basais (pontos de crescimento rentes ao solo), eram mais resistentes à destruição do que árvores. Essa mesma característica torna as gramíneas resistentes ao fogo, ao pastejo e à seca.
Diversificação silenciosa
Com o mundo quente e úmido do Paleoceno e Eoceno, as florestas dominavam. Gramíneas permaneceram componentes minoritários da vegetação por dezenas de milhões de anos, diversificando-se lentamente nos sub-bosques e margens de rios. As subfamílias Bambusoideae e Ehrhartoideae (que inclui o arroz) se diferenciaram nesse período. O registro de fitólitos mostra aumento gradual da presença de gramíneas.
Primeiros campos abertos
O resfriamento global após a glaciação antártica (~33,9 Ma) e a queda nos níveis de CO₂ criaram condições mais secas e sazonais. Florestas recuaram. Pela primeira vez, habitats abertos dominados por gramíneas começaram a aparecer em partes da América do Sul e da África. O registro de fitólitos mostra a transição de vegetação florestal para mista.
A Revolução das Gramíneas
O grande momento. O resfriamento global, a queda do CO₂ atmosférico abaixo de ~500-800 ppm e o aumento da sazonalidade criaram as condições ideais para a explosão das gramíneas C4. Campos abertos substituíram florestas em vastas áreas continentais da África, Américas e Ásia. A fauna se transformou: cavalos evoluíram de animais de floresta com três dedos para pastadores com casco único e dentes hipsodônticos (de coroa alta, resistentes ao desgaste da sílica nas gramíneas). Ruminantes diversificaram-se. Predadores velozes prosperaram nas planícies abertas. Foi a maior reestruturação de ecossistemas terrestres desde a extinção dos dinossauros.
O bioma dominante
Gramíneas cobrem aproximadamente 40% da superfície terrestre (excluindo Groenlândia e Antártica). A família Poaceae possui cerca de 12.000 espécies. Trigo, arroz, milho, sorgo, cana-de-açúcar e centeio são todos gramíneas: elas sustentam a base calórica da civilização humana. As pastagens naturais são hoje consideradas o bioma mais ameaçado do planeta, com mais de 70% já convertidas para agricultura.
Dinossauros comiam grama?
Por décadas, a paleontologia assumiu que gramíneas surgiram somente após os dinossauros. Em 2005, coprólitos de titanossauros na Índia provaram o contrário.
O que são fitólitos?
Fitólitos são estruturas microscópicas de sílica (SiO₂) que se formam dentro das células vegetais. Cada subfamília de gramínea produz fitólitos com formas diagnósticas distintas: bilobados, em sela, em cruz. Quando a planta morre ou é digerida, os fitólitos resistem à decomposição e permanecem no solo, em sedimentos, ou em coprólitos por milhões de anos. São como "impressões digitais" botânicas que sobrevivem ao tempo geológico.
O que encontraram
Nos coprólitos da Formação Lameta (Índia central, ~67 Ma), Prasad et al. identificaram fitólitos de pelo menos cinco linhagens distintas de gramíneas, incluindo representantes das subfamílias Bambusoideae e Ehrhartoideae (a subfamília do arroz). A diversidade encontrada indica que a família já estava bem diferenciada antes da extinção dos dinossauros. Em 2011, um segundo estudo do mesmo grupo, publicado na Nature Communications, demonstrou que a tribo do arroz (Oryzeae) já havia se diversificado no Cretáceo Superior.
Antes vs. depois da descoberta
Visão antiga
"Gramíneas surgiram no Cenozoico, após a extinção dos dinossauros. Nenhum dinossauro jamais comeu grama. Os campos abertos são um fenômeno pós-dinossauros."
Visão atual
"Gramíneas surgiram no Cretáceo (80-100 Ma) e coexistiram com dinossauros por pelo menos 30 milhões de anos. Titanossauros comiam grama. Os campos abertos vieram depois, no Mioceno."

Fotossíntese C4: a arma secreta
A maioria das plantas usa a via fotossintética C3. Algumas gramíneas desenvolveram algo melhor.
O que é a via C4?
Um mecanismo fotossintético modificado que concentra CO₂ ao redor da enzima RuBisCO, reduzindo a fotorrespiração. É mais eficiente que a via C3 em condições de calor intenso, luminosidade alta, seca e baixo CO₂ atmosférico.
Evolução convergente extrema
A fotossíntese C4 evoluiu independentemente pelo menos 22 a 24 vezes dentro das gramíneas, um dos exemplos mais notáveis de convergência evolutiva em plantas. A pressão seletiva da queda do CO₂ no Oligoceno-Mioceno favoreceu esse mecanismo em múltiplas linhagens simultaneamente.
Impacto desproporcional
Gramíneas C4 representam apenas ~3% das espécies de plantas, mas respondem por 23-25% da produtividade primária terrestre global. Dominam savanas tropicais e subtropicais. Milho, cana-de-açúcar, sorgo e milheto são todos C4.
Como as gramíneas mudaram a Terra
Uma família de plantas que redesenhou ecossistemas, solos, regimes de fogo e a evolução dos animais.
Ciclo grama-fogo
Gramíneas produzem biomassa seca, contínua e inflamável que queima facilmente e regenera rapidamente a partir de meristemas subterrâneos. O fogo mata árvores jovens, mas as gramíneas rebrotam em semanas. Isso cria um ciclo de retroalimentação: mais gramíneas geram mais fogo, que impede o avanço das florestas, o que favorece mais gramíneas. A expansão das pastagens no Mioceno coincide com o aumento de depósitos de carvão vegetal no registro sedimentar.
Solos mais férteis do planeta
Gramíneas possuem sistemas radiculares densos e fibrosos que penetram até 2-3 metros de profundidade. Essas raízes contribuem quantidades massivas de carbono orgânico ao solo, criando os solos mais férteis do mundo: os Molissolos (Chernozens). As Grandes Planícies norte-americanas, a estepe ucraniana e os pampas argentinos devem sua fertilidade a milhões de anos de acumulação de matéria orgânica por gramíneas.
Transformação da fauna
A Revolução das Gramíneas no Mioceno reestruturou a fauna de todos os continentes. Cavalos evoluíram de Mesohippus (3 dedos, dentes baixos para folhas) para Equus (casco único, dentes hipsodônticos para gramíneas abrasivas ricas em sílica). Ruminantes como bovídeos e cervídeos diversificaram-se com estômagos de múltiplas câmaras para digerir celulose. Predadores cursórios (corredores) como felinos e canídeos prosperaram nas planícies abertas.
As gramíneas em números
~40%
da superfície terrestre
12.000
espécies conhecidas
100 Ma
fóssil mais antigo
22-24x
vezes que C4 evoluiu independentemente
5+
tipos de gramínea em coprólitos de titanossauros
>70%
das pastagens nativas já perdidas
Referências
Gramíneas
Prasad, V., Strömberg, C.A.E., Alimohammadian, H. & Sahni, A. (2005). Dinosaur coprolites and the early evolution of grasses and grazers. Science, 310(5751), 1177-1180.
Strömberg, C.A.E. (2011). Evolution of grasses and grassland ecosystems. Annual Review of Earth and Planetary Sciences, 39, 517-544.
Cerling, T.E. et al. (1997). Global vegetation change through the Miocene/Pliocene boundary. Nature, 389(6647), 153-158.
Prasad, V. et al. (2011). Late Cretaceous origin of the rice tribe provides evidence for early diversification in Poaceae. Nature Communications, 2: 480.
Florestas mesozoicas e dieta dos dinossauros
Hummel, J. et al. (2008). In vitro digestibility of fern and gymnosperm foliage: implications for sauropod feeding ecology. Proc. R. Soc. B, 275(1638), 1015-1021.
Bomfleur, B., McLoughlin, S. & Vajda, V. (2014). Fossilized nuclei and chromosomes reveal 180 million years of genomic stasis in Royal Ferns. Science, 343(6177), 1376-1377.
Bennettitales
Crane, P.R. (1985). Phylogenetic analysis of seed plants and the origin of angiosperms. Annals of the Missouri Botanical Garden, 72(4): 716-793.
Crepet, W.L. (1974). Investigations of North American cycadeoids: the reproductive biology of Cycadeoidea. Palaeontographica Abt. B, 148: 144-169.
Revolução das angiospermas
Gomez, B. et al. (2015). Montsechia, an ancient aquatic angiosperm. PNAS, 112(35), 10985-10988.
Sun, G. et al. (2002). Archaefructaceae, a new basal angiosperm family. Science, 296(5569), 899-904.
Coevolução flores-polinizadores
Poinar, G.O. & Danforth, B.N. (2006). A fóssil bee from Early Cretaceous Burmese amber. Science, 314(5799), 614.
Peñalver, E. et al. (2012). Thrips pollination of Mesozoic gymnosperms. PNAS, 109(22), 8623-8628.
Fern spike
Tschudy, R.H. et al. (1984). Disruption of the terrestrial plant ecosystem at the Cretaceous-Tertiary boundary. Science, 225(4666), 1030-1032.
Vajda, V. & Bercovici, A. (2014). The global vegetation pattern across the Cretaceous-Paleogene mass extinction interval. Global and Planetary Change, 122, 29-49.